Головной мозг, передний отдел центральной нервной системы позвоночных животных и человека, помещающийся в полости черепа. Г. м.— материальный субстрат высшей нервной деятельности и главный регулятор всех жизненных функций организма.
У беспозвоночных животных, имеющих центральную нервную систему, функцию Г. м. выполняет головной ганглий, настолько развитый у высших насекомых и моллюсков, что его также называют Г. м. Г. м. состоит из конечного мозга (больших полушарий);межуточного мозга, в который входят зрительные бугры (таламус), подбугорье (гипоталамус), забугорье (метаталамус), надбугорье (эпиталамус); среднего мозга, включающего ножки мозга и четверохолмие; заднего мозга, состоящего из моста и мозжечка; продолговатого мозга (рис. 1). Продолговатый мозг — непосредственное продолжение спинного мозга. Все отделы, расположенные между спинным мозгом и межуточным мозгом, образуют ствол мозга. Через него проходят афферентные (центростремительные, чувствительные) нервные волокна, направляющиеся от спинного мозга и черепно-мозговых нервов к вышележащим отделам Г. м., и эфферентные (центробежные, двигательные) нервные волокна, идущие в обратном направлении. Ствол мозга содержит группы специфических афферентных нервных клеток (ядра), воспринимающих информацию от кожных и мышечных рецепторов, расположенных в области головы, а также от других органов чувств (слух, равновесие, вкус). В стволе мозга расположены скопление нервных клеток в виде структуры, называемое сетчатым образованием, или ретикулярной формацией, и ряд нервных центров, ведающих жизненно важными функциями (дыхание, кровообращение, пищеварение и др.).
Примитивный Г. м. имеется уже у предшественника позвоночных животных — ланцетника. В ряду позвоночных Г. м. постепенно усложняется и в нём формируются перечисленные отделы (рис. 2). Постепенное усложнение Г. м. прослеживается во время эмбрионального развития (рис. 3).
Самого высокого развития Г. м. достиг у человека, главным образом за счёт увеличения и усложнения строения двух больших полушарий, морфологически и функционально соединённых мощным пучком нервных волокон — мозолистым телом. В среднем Г. м. взрослого человека весит 1470 г., его объём — 1456 см3, поверхность — 1622 см2. Причём по абсолютным цифрам Г. м. человека уступает только мозгу кита (6000—7000 г) и слона (5700 г). Относительная же масса Г. м., по показателю Я. Я. Рогинского, у человека самая высокая (человек — 32; дельфин— 16; слон — 10,4; обезьяна — 2—4). Увеличение поверхности больших полушарий Г. м. человека и высших животных шло путём нарастания числа борозд и извилин, которые образуют доли полушарий (лобная, теменная, височная, островковая, затылочная и поясная). Большие полушария Г. м. состоят из: 1) поверхностного слоя серого вещества, называемого корой больших полушарий головного мозга; у человека толщина этого слоя 1—5 мм; общее число нейронов в коре около 14 млрд.; их связывают друг с другом и другими отделами Г. м. и спинного мозга афферентные, эфферентные и ассоциативные нервные волокна. В коре, как и в других структурах мозга, имеются глиальные клетки (нейроглия, или глия), которые участвуют в обменных процессах нервной ткани, выполняют опорную функцию и. возможно, играют какую-то специфическую роль в мозговой деятельности; 2) белого вещества, образуемого нервными волокнами, направляющимися в мозг с периферии и идущими от Г. м. на периферию, а также волокнами, связывающими разные участки коры и оба полушария; 3) ряда подкорковых узлов (базальные ганглии), находящихся в глубине полушарий, то есть в толще белого вещества, но состоящих из серого вещества; главнейшие из этих ганглиев —полосатое тело и бледный шар.
Г. м. покрыт твёрдой, паутинной и мягкой мозговыми оболочками, между которыми находится цереброспинальная жидкость, заполняющая также полости мозговых желудочков. Кровеносная система Г. м. и цереброспинальная жидкость служат транспортными руслами питательных веществ, кислорода и других веществ, необходимых для жизнедеятельности нейронов. По этим же руслам из мозга удаляются продукты распада. Г. м. весьма чувствителен к недостатку кислорода.
По ряду анатомичеких и функциональных признаков Г. м. можно представить как совокупность сенсорных систем. Рецепторы (нервные окончания) какой-либо афферентной системы воспринимают раздражения, которые затем в виде нервных импульсов распространяются по центростремительным нервным путям к Г. м. Потоки нервных импульсов несут в Г. м. информацию о силе и качестве раздражений, воспринятых рецепторами органов чувств (глаза, уха, кожи и др.), всех внутренних органов, мышц и сухожилий. В подкорковых структурах, затем в корковых отделах анализаторов, а в конечном итоге всей корой эта информация перерабатывается — осуществляются её анализ и синтез. Затем Г. м. посылает исполнительным органам (эфферентным системам) команды о характере ответных реакций на раздражения. Ответные реакции могут быть двух типов: безусловные рефлексы или условные рефлексы. Двигательные рефлексы осуществляются преимущественно при участии экстрапирамидной системы, состоящей из подкорковых узлов: полосатое тело получает импульсы из таламуса и из коры и передаёт их бледному шару, откуда они поступают в ядра ствола мозга и, наконец, к двигательным нейронам передних рогов спинного мозга. У низших позвоночных (рыб, земноводных и пресмыкающихся) эта система координаций движений единственная. У млекопитающих, кроме неё, появляется пирамидная система, по которой непосредственно передаются импульсы от коры к двигательным нейронам спинного мозга. Она достигает у обезьян и человека высшего уровня развития и обеспечивает наиболее сложные условнорефлекторные, произвольные движения. Пирамидная система, будучи взаимосвязанной с экстрапирамидной, играет уже ведущую роль. Безусловные вегетативные реакции (сосудистые, секреторные, обменные и т. п. ) осуществляются нервными центрами таламуса, гипоталамуса и других структур ствола мозга. Кора больших полушарий связана и с этими структурами, поэтому могут возникать различного рода вегетативные условные реакции (см. Вегетативная нервная система). Нормальная работа Г. м. возможна лишь при определённом уровне возбудимости его основных отделов. Существуют три пути поддержания этого уровня. Первый — через ретикулярную формацию ствола мозга, куда поступают импульсы по ответвлениям (коллатералям) от центростремительных путей, идущих в таламус, а оттуда к соответствующим областям коры. После переработки в ретикулярной формации нервные импульсы утрачивают специфические черты принадлежности к определённому анализатору и приобретают неспецифический характер. Эта импульсация в нужный момент направляется по восходящим путям во все области коры Г. м. и активирует их — задаёт определённый уровень возбудимости (тонус). Второй путь поддержания тонуса коры — через симпатическую нервную систему и мозжечок. Наконец, третий — через специфические пути, идущие от органов чувств. В процессе поддержания тонуса могут принимать участие и условнорефлекторные механизмы. Предполагают наличие у высших позвоночных животных корковой саморегуляции (в том числе и саморегуляции тонуса коры), которая особенно развита у человека. Саморегуляция тонуса обеспечивается двусторонними связями между корой и ретикулярной формацией, а также симпатической нервной системой и мозжечком. Интенсивно исследуются саморегуляторные механизмы Г. м., обеспечивающие те уровни высшей нервной деятельности человека, которые называются мышлением, сознанием и определяются способностью мозга воспринимать, перерабатывать, хранить информацию и выдавать результаты её переработки.
Большую роль в деятельности Г. м. играет лимбическая система, расположенная на внутренней поверхности полушарий Г. м. и в глубине боковых желудочков. Состоит она из гиппокампа, перегородки, миндалевидных тел, грушевидной и поясной извилин, сосковидных тел, бахромы. Иногда включают в её состав также таламус и гипоталамус (и ряд других структур). Предполагают, что лимбическая система имеет отношение к инстинктивным, наследственным реакциям, обусловливающим врожденную основу эмоций, и к некоторым видам памяти. У человека наблюдались расстройства некоторых видов памяти при значительном разрушении гиппокампа и миндалевидных ядер. Пациенты в этих случаях помнят события, предшествовавшие операции, но если их отвлечь чем-нибудь, то они не могут вспомнить, что они намеревались сделать 5—10 мин назад. Разрушение отдельных структур лимбической системы у животных сопровождается нарушением последовательности действий; животное, не завершив одно движение, начинает другое. Электрическое раздражение миндалевидных ядер, перегородки, гипоталамуса у обезьян вызывает драчливость, агрессивность и усиление половой активности. При этом могут меняться взаимоотношения между отдельными особями в стаде: «подчинённая» обезьяна становится «господствующей» и наоборот.
Несмотря на значительные успехи в изучении функции Г. м., в чём наука многим обязана классическим трудам И. М. Сеченова, И. П. Павлова, В. М. Бехтерева, Ч. Шеррингтона, внутренние механизмы его интегративной, целостной деятельности всё ещё остаются невыясненными. В связи с этим строение и функции Г. м. подвергаются интенсивному изучению в лабораториях и клиниках многих стран мира при помощи физиологических, психологических, клинических, биохимических, биофизических, морфологических, кибернетических и других методов исследования.
Лит.: Шмальгаузен И. И., Основы сравнительной анатомии позвоночных животных, 4 изд., М., 1947, с. 225—76; Орбели Л. А., Вопросы высшей нервной деятельности, М. — Л., 1949, с. 397—419, 448—63; Павлов И. П., Полн. собр. соч., т. 3, кн. 2, М. — Л., 1951, с. 320—44; Быков К. М., Кора головного мозга и внутренние органы, Избр. произв., т. 2, М., 1954, с. 358—84; Сеченов И. М., Рефлексы головного мозга, М., 1961; Воронин Л. Г., Курс лекций по физиологии высшей нервной деятельности, М., 1965, с. 225—59; Физиология человека, М., 1966, гл. 15; Проссер Л., Браун Ф., Сравнительная физиология животных, пер. с англ., М., 1967, гл. 21; Лурия А. Р., Высшие корковые функции человека..., М., 1969, с. 7—80.
Л. Г. Воронин.

Рис. 2. Постепенное усложнение головного мозга у позвоночных животных (вид мозга сверху): А — мозг акулы; Б — лягушки; В — аллигатора; мозг млекопитающих: Г — тупайи; Д — лошади; Е — человека (вид сбоку). 1 — обонятельная доля; 2 — обонятельная луковица; 3 — эпифиз; 4 — третий желудочек; 5 — зрительная доля; 6 — мозжечок; 7 — продолговатый мозг; 8 — межуточный мозг; 9 — четвёртый желудочек; 10 — большие полушария; 11 — извилина; 12 — борозда.
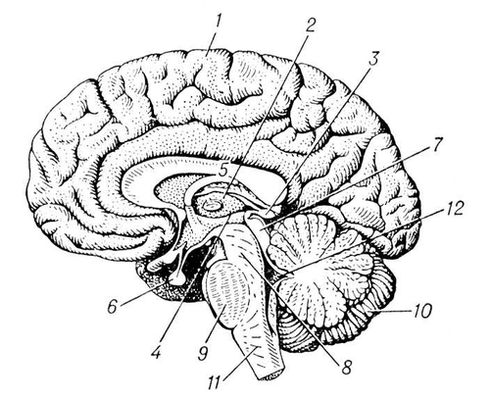
Рис. 1. Головной мозг взрослого человека (правая половина, вид слева): 1 — большое полушарие; 2 — зрительный бугор (таламус); 3 — надбугорье (эпиталамус); 4 — подбугорье (гипоталамус); 5 — мозолистое тело; 6 — гипофиз; 7 — четверохолмие; 8 — ножки мозга; 9 — мост (варолиев); 10 — мозжечок; 11 — продолговатый мозг; 12 — четвёртый желудочек.

Рис. 3. Боковая поверхность мозга человека на различных стадиях эмбрионального развития (конечный мозг заштрихован): в возрасте 2 недель (1), 3 недель (2), 4 недель (3), 8 недель (4), 6 месяцев (5); мозг взрослого человека (6).