Различными исследователями могут быть учтены те или иные признаки и их вариации, и при этом практически для идентичных признаков "бессознательно" использованы совершенно иные системы их обозначения (кодирование фенов). Это в значительной мере затрудняет сопоставление данных разных авторов и препятствует рациональному и эффективному накоплению и "сбережению" экспериментального материала. Определенные трудности возникают при обработке данных фенетических исследований, необходимости подробного описания анализируемых признаков и целесообразности опубликования материалов явно описательного характера.
Единственным правильным решением, устраняющим эти "несуразности", является составление каталогов фенов и фенокомплексов для изучения тех или иных видов древесных пород. В настоящее время, к сожалению, имеется весьма скудный и ограниченный материал по этому разделу фенетики популяций древесных пород, однако постепенно экспериментальный материал по основным лесообразующим породам существенно пополняется, и в ближайшем будущем уже можно будет приступить к объединению и разработке единого каталога фенов хотя бы основных лесообразователей страны. В принципе уже сейчас нужно немедленно объединить весь накопившийся материал различных авторов, работающих в этом "ключе" с древесными растениями, но при этом надо иметь в виду, что каталогизации древесных растений должна предварительно предшествовать разработка "единого типа записи" фенетических признаков (система кодирования).
Последующим этапом должна быть организация центра, где бы концентрировались все материалы (как опубликованные, так и рукописные), имеющие отношения к данному разделу исследований (к примеру, по лесным древесным породам – ФГУП "Научно-исследовательский институт лесной генетики и селекции"), причем чтобы каждое заинтересованное лицо в любой момент смогло бы ими воспользоваться.
Завершение процедуры каталогизации фенов существенно "продвинет" вперед фенетические исследования популяций древесных растений. В перспективе необходимо расширить спектр исходных признаков и приступить к составлению каталога атласа фенов в пределах ареалов каждой из лесообразующих пород, разработать теоретические аспекты фенетики популяций древесных растений с учетом специфики их видового разнообразия.
2.3. СОСНА ОБЫКНОВЕННАЯ (P1NUS SYLVESTTUS L.)
КАК УНИВЕРСАЛЬНЫЙ МОДЕЛЬНЫЙ ОБЪЕКТ ДЛЯ ФЕНЕТИЧЕСКИХ ИССЛЕДОВАНИЙ
Сосна обыкновенная как полиморфный вид в плане изучения на фенетическом уровне представляет исключительный интерес в качестве объекта исследований. Род сосен (Pinus) насчитывает более 100 видов, произрастающих в основном в лесной зоне Евразии и Северной Америки. На территории бывшего СССР находится 14 дикорастущих видов, занимающих по площади и запасам второе место после лиственницы и относящихся к двухвойным и пятихвойным соснам. Из двухвойных сосен наиболее распространена здесь сосна обыкновенная или лесная (Pinus sylvestris L.), и в связи с большой индивидуальной изменчивостью и огромным ареалом этот вид имеет множество морфологических и экологических форм и разновидностей.
По биологии, экологии и систематике рода Pinus и сосны обыкновенной основной перечень работ может быть сведен к следующему: С.З. Курдиани (1910), Э.Л. Вольф (1925), В.Н.Сукачев (1928, 1938), С. С. Голубовский (1934), Г. Д. Гальперн (1949), А. Н. Юновидов (1950), О. Г. Каппер (1954), А. М. Семенова-Тян-Шанская (1956), В. Б. Сочава (1956), А. П. Шиманюк (1962), Л. Ф. Правдин (1964), Ф. Н. Харитонович (1968), Леса СССР (1966 – 1970), С. А. Мамаев (1972), Н. В. Кречетова (1974), Л. П. Рысин (1975), А. В. Побединский (1979), Атлас лесов СССР (1973), Древесные породы мира, Т. 3: Древесные породы СССР (1982), М. М. Котов (1982), В. К. Мякушко и др. (1989), Г. Ф. Морозов (1922, 1924) А. Д. Букштынов и др. (1981), И. А. Алексеев (1969), А И. Чернодубов (1996), М.Н.Егоров (1997, 2001) и др. По последним данным, сосна обыкновенная занимает около 16 % земли Гослесфонда, и из имеющихся сейчас в стране лесных культур на ее долю приходится 34 %. Накопившийся к данному моменту перечень публикаций по внутривидовой изменчивости сосны обыкновенной весьма обширен, и обзор его является актуальным и необходимым для раскрытия ее полиморфизма и биологической сущности.
В частности, впервые "пирамидальная сосна" была описана у нас в начале XX столетия неким Б., по habitus'u резко отличающаяся от окружающих сосен (Барышевцев В. В., 1908). Позднее мы встречаем ее описание у М. П. Скрябина (1951) и С. А. Петрова (1961).
Широко- и узкокронные формы сосны описывает Е. Zedeibauer (1912), Б. Д. Жилкин (1960) и Л. С. Василевская (1962) в лесах Белорусской ССР, Н. П. Мишуков (1964), Е. Г. Орленко (1965) и М. Н. Егоров (1981).
Деление сосны на расы приводят С. 3. Курдиани (19086, 1913, 1916), Н. Н. Кобранов (1914), Д. И. Морохин (1923), С. А. Самофал (1925) и П. Я. Соколов (1928).
Наличие форм сосны в зависимости от цвета женских и мужских стробилов устанавливают Т. П. Некрасова (1959), Г. М. Козубов (1962а), С. А. Мамаев (1965, 1972), Е.А. Пугач (1976, 1982), К. А. Кузьмина (1978), М. Н. Егоров (1983), Л. Ф. Правдин (1950). Ю. Н. Азниев (1979) описывает половой диморфизм у сосны обыкновенной. Трехпучковую сосну отмечают в своей сводке М М. Вересин и М. К. Улюкина (1979). Описание «воротничковой» коры у сосны находим у Б. В. Гроздова (1958) и А. А. Божок (1976).
Е. С. Петренко (1959), А. А. Хиров (1973) и В. В. Шульга (1979) дают характеристику сосны карликовой и с "ведьминои метлой". Феномен гроздешишечной сосны мы встречаем у Б. Ожгутова (I960), Е. П. Проказина и Т. П. Проказиной (1962), А. А. Листова (1971) и М. М. Вересина (1979).
Изменчивость морфологических признаков шишек и семян сосны детально обсуждается в работах А. Н. Соболева (1908), В. П. Князева (1954), С. А. Мамаева (1959) для сосны Мещерской низменности, И. Н. Лигачева (1959, 1962) для сосняков Бурятской АССР, В. Л. Черепнина (1970), В. В. Шульги (1972, 1973) для сосны из Казахстанских островных боров, В. П. Бутаева (1976) для сосны из Якутии, А. В. Хохрина (1977, 1981), А. М. Голикова (1979, 1981), Г. С. Захаренко (1986), А. Н. Толстопятенко (1989, 1991), А. И. Видякина (1991) для популяций сосны на востоке европейской части России, М. Н. Егорова (1976, 1993) для сосны из Пермской, Кировской, Свердловской и Крымской областей, А. И. Чернодубова (1996) для островных боров Восточно-Европейской равнины.
Индивидуальная и экологическая изменчивость сосны анализируется в трудах В. И. Носкова (1954), Е. Г. Орленко (1965, 1973), Д. А. Стецкой (1960), Г. М. Козубова (19626) для сосны из Карелии, Н. П. Мишукова (1966) в Новосибирской области, И. П. Положенцева и М. Э. Муратова (1968) на Южном Урале, С. А. Мамаева (1972, 1974) и А. А. Хирова (1973) в Бузу- лукском бору, В. М. Долгошеева (1974), И. И. Бауманис и др. (1975), Е. П. Проказина и Л. А. Бопп (1976), М. С. Ковалева и Л. С. Ковалева (1986), М. Н. Егорова (1972 – 1994).
Формовое разнообразие сосны по ряду структурных признаков нашло свое отражение в публикациях В. Н. Гаврись (1938), А. В. Альбенского (1941) в Забайкалье, Г. Ф. Привалова (1960) в лесной опытной даче СО АН СССР, Н. В. Когеловой (1961), Л. Ф. Правдина (1964), С. А. Мамаева (1965), В. А. Николаюка (1972) да Брянской области, А. И. Савченко и др. (1972) для лесов Белоруссии, Ю. И. Азниева (1976), А. М. Голикова (1978) для Псковской области, В. В. Грицайчук (1979) для ленточных боров Западной Сибири, В. В. Удилова (1990) для популяции сосны Среднего Урала.
Тем не менее с учетом накопившегося и рассмотренного выше фактического материала по внутривидовой изменчивости сосны обыкновенной и анализа особенностей методических подходов разных авторов к изучению изменчивости нами применительно к лесным древесным растениям был предложен целый ряд новых методических решений, основанных на использовании комплекса признаков как качественной, так и количественной природы их происхождения. Набор ведущих признаков с качественной (неметрической) и количественной (размерной) природой их происхождения, естественно, может быть расширен или сужен в зависимости от поставленной задачи в эксперименте и их хозяйственной ценности.
При этом всегда нужно помнить, что число признаков практически бесконечно, если к тому же еще "раздвинуть" круг структурных признаков подключением к ним функциональных и биохимических; и чем представительнее будет их набор, тем ближе осуществляется подход к установлению аллелофонда как суммы всех аллелей особи, вида и популяции в целом.
Из приведенных данных видно, что сосна обыкновенная как модельный объект для фенетических исследований представляет большой интерес для использования ее структурных и физиолого-биохимических характеристик в селекционном анализе.
ГЛАВА 3
ФЕНОФОНД ДРЕВЕСНЫХ РАСТЕНИЙ И ЕГО ДИНАМИКА
3.1. ФЕНОФОНД
Фенофондом называется все многообразие фенов в пределах какой-либо группы особей. По аналогии с понятием гено (аллело) фонда можно говорить о фенофонде популяции, групп популяций, отдельных демов и т.п. (Яблоков А. В., 1978). Изучение фенофонда, как правило, включает в себя его выявление, методы описания, динамику и пути сравнения фенофондов. Обычно фенофонд исследуют в двух направлениях: изучение популяционной статики и популяционной динамики. В первом случае выявляются внутрипопуляционные структуры, так как фенами маркируются любые внутрипопуляционные группировки; во втором случае вскрывается ход процесса микроэволюции, а также выделяется и определяется степень давления отдельных эволюционных факторов на действие "пусковых механизмов эволюции".
К настоящему моменту пока нет достаточно полного описания фенофонда какого-либо вида живых организмов, включающего несколько тысяч фенов, имеющих отношение ко всем типам признаков (морфологических, физиологических и биохимических) и отражающих если не все множество генотипов, то хотя бы значительную часть. Предстоит большая и кропотливая работа, чтобы в пределах ареала каждой из лесообразующих пород дать подробную информацию о фенофонде и фенотипической структуре той или иной древесной породы, чтобы иметь более или менее полное представление о частотах встречаемости маркерных признаков-фенов и их миграционном дрейфе из одной популяции в другую в границах ареалов изучаемых видов. Методы описания фенофондов могут быть самые различные – это буквенное и числовое кодирование фенов при описании фенофонда, графическое его представление (гистограммы, различные секториальные графики или векторные диаграммы и др.).
При оценке данных могут быть использованы методы хи-квадрат, критерий Стьюдента, ранговый коэффициент корреляции Спирмена, формула Харди-Вайнберга, коэффициент дистанции Массатоши Неи (Nei М., 1972), координатный метод комплексной оценки лесных древесных пород и методы сравнительного многомерного анализа (СМА) – метод главных компонент, таксономия, распознавание образов, дискриминантный, факторный и кластерный анализы. Не исключается и использование балльного сравнения, где вариации отдельного признака выражаются определенным числом баллов, и сумма их по всем изучаемым признакам дает обобщенную характеристику особи.
Со времени проведения I Всесоюзного совещания-семинара по фенетике популяций (Саратов, 1976) уже накоплен определенный опыт путей и методов изучения фенофондов основных лесообразующих пород страны.
Л. Ф. Правдин (1964) при анализе фенофонда древесных растений предложил ряд методических решений – это методики изучения изменчивости морфолого-анатомических, эколого-физиологических признаков в зависимости от широты и долготы их местопроизрастания.
Н. В. Тимофеев-Ресовский и А. В. Яблоков (1973) констатируют: "Сегодня можно говорить о нескольких крупных направлениях в феноанализе. Это – популяционная морфофизиология, феногеография и изучение фенофонда" (с. 45) и "в качестве еще одного направления популяционной морфофизиологии назовем установление степени сходства и различия между отдельными популяциями" (с. 48).
С. А. Мамаев и А. К. Махнев (1982, 1983, 1988) при изучении фенофонда предлагают поэтапную оценку форм изменчивости древесных растений (эндогенная, сезонная, экологическая, индивидуальная и географическая) с подключением комплексного метода морфофизиологических маркеров с подразделением их на эволюционно относительно стабильные и лабильные.
Л. И. Милютин (1982–1990) при анализе экспериментального материала рекомендует широко применять метод средних гибридных индексов (обобщенное балльное сравнение фенофондов), математические методы многомерной классификации, дискриминантный и кластерный анализы, расстояние Мехалонобиса.
М. В. Придня (1985) указывает, что по дифференциации семей и популяций кавказской пихты на цельнолистный и выямчатолистный фенотипы формируется прообраз фитоценоза, в котором первые составляют доминирующий ярус, вторые – подчиненный.
А И Ирошников (1983, 1985) при изучении фенофонда природных популяций для ряда случаев обращает внимание на постановку экспериментов по испытанию определенных генотипов на разном экологическом фоне и исследованию их пространственной структуры (мозаичность распределения фенофонда, обусловленная неоднородностью среды и ограничениями панмиксии).
Л. Ф. Семериков и Н. В. Глотов (1983) с помощью иерархического дисперсионного анализа для характеристики пространственной структуры вида и выяснения причин дифференциации популяций вычисляют оценки обобщенных различий между субпопуляциями с применением методов распознавания образов, разработанных в ИММ УНЦ СССР (Семериков Л. Ф. и Казанцев Н. П., 1979).
Б. А. Кравцов и Р. Г. Хлебопрос (1985), используя методы нумерической таксономии в фенетике популяций, на примере лиственниц Сибири показали полезность ряда непараметрических процедур при фенетическом анализе природных популяций, где наглядно продемонстрировано преимущество одновременного изучения фенов по сравнению с обычно используемыми методами, основанными на последовательном их рассмотрении. Предложена методика ранжирования фенов по их информационной ценности (Кравцов Б. А., Милютин Л. И., 1981, 1983).
М. Н. Егоров (1983 – 2001) предусматривает в программе изучения фенофонда выявление структуры и динамики фенотипического состава популяций, описание и сравнение их фенофондов на разных фазах онтогенеза внутри одного поколения и разных поколений, а также в условиях изоляции. При оценке данных могут быть использованы СМА и координатный метод комплексной оценки лесных древесных пород.
С. А. Петров (1985, 1990), обобщая имеющиеся опубликованные материалы по фенотипической структуре популяций видов лесных древесных растений, а также используя результаты собственных исследований, приходит к следующим заключениям: распределение частот различных фенов в популяциях подчиняется распределению Харди-Вайнберга; концентрация отдельных фенов в популяциях подвержена клинальной изменчивости в различных направлениях – долготном и широтном, а также высотной поясности; в малых по объему популяциях концентрация отдельных фенов резко повышается. Краевые популяции характеризуются более высоким уровнем полиморфизма по сравнению с центральными, и изменчивость популяций по многим фенам подчиняется закону гомологических рядов в наследственной изменчивости Н. И. Вавилова (1920).
В. М. Максимов и И. В. Кошовской (1990), изучая фенотипическую структуру популяций сосны обыкновенной в ЦЧО по составу эфирных масел, пришли к выводу, что для сосны Усманского бора характерны три основных типа биосинтеза монотерпенов: каренистый, промежуточный и пиненистый и наблюдается связь с частотой встречаемости каренистого и пиненистого типов биосинтеза эфирного масла хвои в зависимости от условий местопроизрастания.
А. И. Чернодубов (1990) на примере изучения структуры популяций сосны обыкновенной по содержанию ?3-карена в эфирных маслах из хвои показал, что по мере продвижения с юго-запада на север уменьшается количество деревьев, продуцирующих ?3-карен, и почти отсутствуют они в северных (Мурманская и Архангельская области) частях ареала.
Как видно из приведенного перечня исследований по фенофонду древесных растений, пути его изучения и применяемые при этом различные методологические подходы могут быть самыми разнообразными, но предусматривающими одну цель – как можно полнее раскрыть его структуру для выявления перспективных, хозяйственно-ценных особей популяционных группировок и использования их в лесохозяйственном производстве.
3.2. ДИНАМИКА ФЕНОФОНДА
Динамика фенофонда является, в свою очередь, неотъемлемой составной частью фенофонда основных лесообразующих пород. "Изучение динамики частот фенов, даже общее для многих популяций вида, выявляет особенности совокупностей различного ранга. Один из возможных путей изучения действия естественного отбора на фенофонд открывается при выяснении определенного адаптивного значения того или иного фена" (Яблоков А. В., Ларина Н. И., 1985, с. 108).
При выявлении особенностей интенсивности элементарных эволюционных факторов фенетика может быть весьма востребованной Первый путь заключается в выполнении действия естественного отбора путем сравнения концентрации тех или иных Ленов в разные моменты жизни одного и того же поколения. Если индивидуум родится с каким-то признаком, который в дальнейшем в течение онтогенеза не изменяется (к примеру, цвет хвои, коры, зрелых шишек, семян, женских соцветий, пыльников, крыла семени, строение коры, форма апофиза шишек, архитектоника кроны и ряд других у сосны обыкновенной), то есть возможность сравнить распределение того или иного признака у разных особей на разных стадиях онтогенеза. В ряде случаев наблюдается заметный сдвиг в частоте встречаемости признака, и по таким сдвигам можно проследить действие естественного отбора.
Другим путем изучения действия отбора является сопоставление фенофонда разных поколений. "Колебания концентрации редких фенов и устойчивость концентрации массовых фенов указывают на существование стабилизирующего отбора по главным признакам и свойствам" (Яблоков А. В., 1980, с. 77). Разумеется, не во всех случаях исследователь сталкивается с постоянством фенотипа на протяжении нескольких поколений (случается, что частота почти всех фенов резко варьирует, и объяснением столь значительных колебаний в частоте встречаемости фенов в смежных поколениях может быть слабое давление отбора на изучаемые признаки или непредставительная выборка для характеристики всей популяции).
Следующим эволюционным фактором, влияние которого можно выяснить посредством изучения фенофонда, является изоляция. Различия в концентрации отдельных фенов позволяют предполагать действие одной из форм изоляции, которых в природе много (территориально-механическая, экологическая, фенологическая и др.). Не последнее место в анализе фенофонда занимает и один из элементарных эволюционных факторов – "волн жизни".
Следовательно, изучение фенофонда дает интересные, а подчас и уникальные возможности исследования трех элементарных эволюционных факторов – естественного отбора, изоляции и волн жизни .
ГЛАВА 4 ФЕНОТИПИЧЕСКАЯ СТРУКТУРА ПОПУЛЯЦИЙ ДРЕВЕСНЫХ РАСТЕНИЙ
4.1. ИЗУЧЕНИЕ СТРУКТУРЫ
Как естественно-исторический феномен живой природы любая популяция обладает только ей присущими особенностями, и прежде всего – структурированностью. Генетическая и фенотипическая структуры популяции могут быть охарактеризованы тем или иным количественным соотношением генотипов и фенотипов.
В нашей стране изучение структуры популяций древесных растений началось с работ В. А. Драгавцева и В. М. Остриковой (1972), В. А. Драгавцева (1962 – 1994), С. А. Петрова (1964, 1970), Е. П. Проказина и А. В. Чудного (1968), Л.Ф.Правдина (1964, 1965, 1967), С. А. Мамаева (1968, 1969), А. И. Ирошникова (1964, 1985).
Обширные исследования популяционной структуры видов дуба европейской части бывшего СССР и Кавказа осуществляет Л.Ф.Семериков (1971, 1975, 1986) и в соавторстве с Н. В. Глотовым (1971, 1980); А. И. Чернодубов (1996), М. Н. Егоров (1997) – по сосне обыкновенной.
С. А. Петров (1974, 1975) отмечает, что, несмотря на успехи, достигнутые в изучении структуры популяций многих видов, до настоящего времени недостаточно исследованы генетические закономерности в популяциях лесных древесных растений. Не выяснен уровень генетического разнообразия популяций по тем или иным признакам и особенно слабо изучена генетика количественных признаков, наиболее сложных и в то же время наиболее ценных с хозяйственной точки зрения. Лесная генетика все еще значительно отстает в изучении одного из главных вопросов – структуры популяций, хотя острая необходимость проведения таких исследований очевидна (Sokolowski S., 1931; Драгавцев В. А., 1962). По данным А. И. Ирошникова (1974), большинству видов древесных растений свойствен значительный полиформизм и в связи с этим изучение генотипического состава и выявление структуры популяций важны для познания направления действия естественного отбора и условий, обеспечивающих устойчивость популяций, принципов оценки особей в целях селекции. При этом появляется возможность более глубокого познания эволюции и объективного выделения внутривидовых категорий.
Л. Ф. Семериков (1986) своевременно подмечает, что стратегия лесной генетики и селекции должна строиться не только на поиске изучении и введении в культуру лучших генотипов, но и на познании механизмов поддержания устойчивости и высокой продуктивности природных популяций, оценке внутривидовой изменчивости и анализе эколого-генетической структуры природных популяций лесообразующих пород в различной биогеоценотической обстановке. Другими словами, речь идет о круге проблем, определяемых как популяционная структура вида.
Актуальность и необходимость изучения фенофондов и фенотипической структуры древесных растений, состояния и перспектив развития исследований в этом направлении были подчеркнуты и нами (Егоров М. Н., 1981 – 2001).
Важность изучения фенофондов и фенотипической структуры природных и искусственных популяций древесных пород продиктована как научными, так и в гораздо большей степени производственными запросами интенсивно развивающегося лесного хозяйства страны. В частности, практическое преломление результатов исследований нацелено на разрешение задач по научно обоснованному формированию качественного состава и продуктивности культур из хозяйственно ценным форм.
К изучению популяций, представляющих собой первый надиндивидуальный уровень интеграции жизни и элементарную эволюционную единицу, чаще привлекаются такие разделы популяционной биологии (хотя провести четкие границы между ними очень трудно), как генетика, экология, морфология, физиология и биология индивидуального развития. В данной работе, в частности, большое место отведено находящейся в стадии активного формирования популяционной морфологии, возникшей как самостоятельное направление в 70-е гг. XX в. и включающей в круг своих задач изучение структуры, состава и динамики популяций посредством изучения морфологических признаков особей, т.е. любых поддающихся сравнению структурных особенностей индивидуума. Наибольший интерес для морфологии представляют сравнительно быстро изменяющиеся качественные и количественные структурные признаки. Популяционная морфология часто трактуется как направление в изучении микроэволюции ввиду проникновения популяционного начала в эволюционную морфологию и ее тесной связи с синтетической теорией эволюции.
Из перечисленных сведений по фенотипической структуре популяций древесных пород видно, что хотя они и получили должное развитие и направление, однако крайне недостаточны, и здесь необходимо приложить еще много усилий, чтобы получить более или менее полную научную информацию для ее последующего обобщения и анализа.
4.2. МОДЕЛИ ФЕНОТИПИЧЕСКОЙ СТРУКТУРЫ ПРИРОДНЫХ И ИСКУССТВЕННЫХ ПОПУЛЯЦИЙ
В настоящее время число различных моделей, использующих самый разнообразный математический аппарат в проблеме математического моделирования биологических сообществ и организмов, непрерывно возрастает; причем отчетливо выделяются два класса моделей – модели природных и искусственных сообществ. Тем не менее специфические особенности биологических систем, такие как сложность строения каждой особи, полиморфность условий среды, пространственная незамкнутость как в энергетическом, так и в структурном смысле, и обширный спектр внешних характеристик, при которых сохраняется жизнеспособность систем, создают определенные сложности в их исследовании и моделировании. При этом правильное выявление направляющего фактора (или группы их) эволюции является необходимым условием для понимания путей в прогнозировании ее хода, а также для управления эволюционным процессом в естественных и особенно искусственно созданных биогеоценозах.
И. И. Шмальгаузен (1935) в свое время говорил о принципиальной невозможности выразить весь ход роста одной математической формулой. К такому же выводу приходит и F. J. Richards (1969).
Математическое моделирование как метод научного исследования нашло применение в первую очередь в популяционной генетике и экологии с основополагающих работ R. А. Fishef (1930), J. В. S. Haldane (1932) и S. Wright (1931). Более детальную разработку методологии системного анализа провелй А. А. Ляпунов и С. В. Яблонский (1963). Интерпретацию общих системных представлений для задач биогеоценологии осуществляет В. Д. Александрова (1964). Ю. Г. Пузаченко (1971) ряд методологических подходов использовал при обосновании методов исследования организации биосистем. А. А. Ляпунов (1975) указывает, что биология в широком смысле, в противовес физике, органически связана с дискретной математикой.
Ю. Г. Пузаченко и B. C. Скулкин (1981) акцентируют внимание на том, что выбор конкретного математического аппарата обеспечивающего синтез модели системы, прямо зависит от конкретных результатов исследования и не может быть определен заранее, и эффективность избранного метода должна проверяться по независимой выборке наблюдений за поведением реальной системы.
Л. Г. Косолапова и Б. Г. Ковров (1988) в математическом моделировании генетических, экологических и эволюционных процессов выделяют два подхода: собственно математическое моделирование как метод теоретической биологии и имитационное моделирование, направленное, главным образом, на решение практических задач.
Тем не менее моделирование как процесс создания модели в современных исследованиях возведен в ранг обязательного этапа любого научного поиска и является "инструментарием" с большой разрешающей силой (Ляпунов А. А. и др., 1963; Александрова В. Д., 1964; Пузаченко Ю. Г., 1971; Maynard Smith J., 1974; Ляпунов А. А., 1975; Свирежев Ю. М., 1975, 1981; Кондаков Н. И., 1976; Джефферс Дж., 1981; Миркин Б. М. и Розен- берг Г. е., 1983; Реймерс Н. Ф., 1991). Для иллюстрации в работе приводится ряд вариантов разработанных моделей, в частности: "TAKS 2.1", "TAKSRS.BAS", "WRMAT 2.BAS", "KLQ", "TAKSRSLBAS", "TAKSRS2.BAS" и др. Сравнение фенофондов и фенотипических структур природных сосняков и культур на основе новых математических моделей осуществлено в работе по таксономическому методу СМА – в программе "TAKS 2.1", по изотропному методу СМА – "TAKSRS2.BAS", кластерному анализу - "KLQ". Координатный метод комплексной оценки и показатели популяционной изменчивости по полиморфным признакам сосны реализованы в соответствии с алгоритмическими схемами данных методов и их программ, приведенных в соответствующих разделах.
4.2.1. Таксономический метод сравнительного многомерного анализа
Классификация как распределение объектов (для нашего случая деревьев) по более или менее однородным группам и установление соотношений между группами прежде всего нашла отражение в работах таксономистов (название происходит от двух греческих слов: таксис – расположение, порядок и номос – правило, принцип, закон). Таксономия как научный подход и метод далеко уходит своими корнями в историческое прошлое (Аристотель, 384 – 322 гг. до н.э.; Теофаст, 370 – 285 гг. до н.э.; Диоскорид и Плиний Старший, 23 – 79 гг.). Однако лишь Карл Линней в XVIII в. разработал иерархическую систему таксономических категорий. Окончательное закрепление бинарной номенклатуры связано с деятельностью О. П. Декандоля (1778 – 1841) и его последователей.
В последние годы на Международных ботанических конгрессах, начиная с IV (Париж, 1967), происходили усовершенствования и уточнения ботанической номенклатуры (Алексеев Е. Б. и др., 1989).
В настоящее время наиболее важным достижением в развитии таксономической методологии было введение в нее филогенетической систематики, кладистики (от слова "клад" – происхождение от одного предка) и появляется соответственно кладистическая классификация. При этом внедрение кладистики Хеннига сопровождалось более ожесточенной дискуссией, чем при внедрении числовой таксономии (Janvier Philippe, 1990). Сейчас четко обозначились контуры трех конкурирующих подходов в систематике – фенетики, кладизма и традистики (Расницын А. П., 1992). Много общих элементов у классической типологии числовой фенетики, кладистики и эволюционной систематики отмечает Л. Н. Васильева (1989). Первое в нашей стране обобщение кла- дистического анализа приводит И. Я. Павлинов (1989).
Нами применительно к древесным породам (к сосне обыкновенной, в частности) предпринята попытка применения таксономического метода СМА к изучению фенофонда и фенотипических структур природных популяций и культур сосны на основе разработанного автором совместно с Л. С. Рыбаковой пакета прикладных программ ОТЭКС для ЭВМ с учетом методических подходов ряда авторов (Плюта В., 1980, 1989; Кашик С. А. и Мазилов В. Н., 1984; Шорников Б. С., 1979).
Одним из первых методов исследования многомерных объектов был предложен таксономический показатель уровня развития di, с помощью которого представляется возможность оценить достигнутый в некоторый период или момент "средний" уровень значения признаков, характеризующих изучаемое явление. Интерпретируется он следующим образом: данная единица тем более развита, чем ближе значение di к единице (Hellwig Z., 1965).
В соответствии с процедурой нахождения di и составленной программой для ЭВМ проведен анализ пробных площадей сосны до комплексу признаков, заложенных в спелых сформировавшихся природных древостоях и одновозрастных с ними культурах сосны 1861 и 1864 гг. производства, идентичных по почвенно-грунтовым условиям местопроизрастания и принадлежащих к одному естественному ряду их роста и развития из лесной (Средний Урал) и лесостепной (Центральное Черноземье) зон. На примере пробных площадей (пр. пл.) № 5, 6 и № 11, 12 приводятся итоги анализа сравниваемых сосняков и особенности их сходств и различий по фенотипической структуре, в частности:
– для 130-летних природных древостоев и культур 1864 г. (Средний Урал, пр. пл. № 5 и 6) процентное соотношение по 9 классовым интервалам значений di, составило соответственно 1 – 8,2 и 6,5; 2 - 0,8 и 1,2; 3 – 16,4 и 14,0; 4 - 11,5 и 17,5; 5 – 22,9 и 17,5; 6 – 14,8 и 12,5; 7 – 6,6 и 11,5; 8 – 8,2 и 6,5; 9 – 11,6 и 0;
– для спелых 130-летних природных древостоев и культур 1861 г. (Центральное Черноземье, пр. пл. № И и 12) процентное соотношение по 8 классовым интервалам значений df 1 – 7,5 и 7,0; 2 – 9,0 и 17,5; 3 – 26,0 и 25,5; 4 – 21,5 и 30,5; 5 – 23,5 и 7,5; 6 – 11,0 и 5,5; 7 – 1,5 и 5,0; 8 – 0 и 1,5.
Таким образом весь цифровой материал указывает на очевидность и доказуемость различий по рассматриваемым классовым интервалам показателя di и практически не нуждается в каких- либо комментариях. В проанализированных парах пробных площадей как по первоначальному, так и по конечному на выходе значению показателя di видно, что культуры не только не уступают, но и превосходят естественные популяции сосны. При сравнении же одновозрастных спелых природных древостоев и культур сосны по изменчивости показателя di из различных условий местопроизрастания (Средний Урал и Центральное Черноземье) просматривается, что преимущество следует отдать сосне из Центрально-Черноземной популяции.
Различие фенофондов природных и искусственных спелых сосновых насаждений наглядно иллюстрируют секториальные графики (векторные диаграммы) – рис. 4.1. и 4.2.

Рис. 4.1. Секториальные графики (векторные диаграммы) для изображения фенофондов природных (Е) и искусственных (К) насаждений сосны VII класса возраста по показателю уровня развития деревьев di (пр. пл. № 5 и 6, Свердловская область, Билимбаевский лесхоз, Первоуральское лесничество, кв. 52)
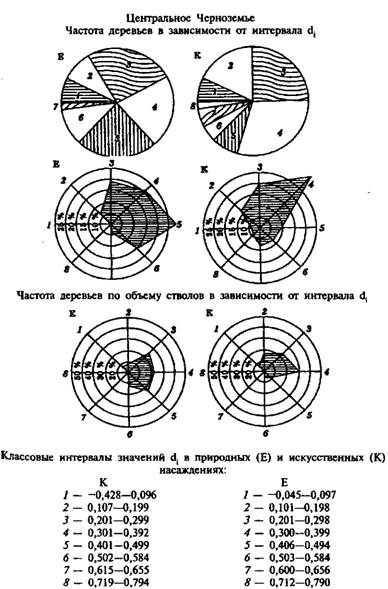
4.2. Секториальные графики (векторные диаграммы) для изображения фенофондов природных (Е) и искуственных (К) насаждений сосны класса возраста по показателю уровня развития деревьев dj (пр. пл. № 11 и 12, воронежская область, Воронежский лесхоз, Углянское лесничество, кв. 46)
В целом можно отметить, что рассмотренный таксономический метод СМА является одним из уникальных с большой разрешающей способностью методологических подходов к изучению фенотипической структуры основных лесообразователей лесного фонда страны и может быть рекомендован к широкому применению в подобного рода исследованиях.
4.2.2. Кластерный анализ
"Как научное направление кластер-анализ заявил о себе в середине 60-х годов и с тех пор бурно развивается, являясь одной из ветвей наиболее интенсивного роста статистической науки" (Мандель И. Д., 1988, с. 4). Кластер-анализ – это способ группировки многомерных объектов, основанный на представлении результатов отдельных наблюдений точками подходящего геометрического пространства с последующим выделением групп как "сгустков" этих точек, и он органически вписывается в общую теорию классификации, которая в последние годы интенсивно развивается (Розова С. Е., 1986). Наиболее ярко и выпукло кластерный анализ отражает черты СМА в классификации объектов, и основные методические черты его сводятся к двум: образование единой меры, охватывающей ряд признаков, и количественное решение вопроса о группировке объектов наблюдения. Классификация лесных сообществ по комплексу структурных морфологических), функциональных (физиологических) и биохимических признаков является одной из важнейших задач формационного и ассоциативного анализов.
В качестве меры группового сходства может быть взята величина арккосинуса коэффициента корреляции между объектами, если объектами выступают изучаемые переменные (R-метод), или величина косинуса угла между векторами, компонентами которых являются измеренные признаки (Q-метод). В основу программы кластерного анализа положены алгоритмы, описанные в работах У. Крамбейна и др. (1973) и Дж. Девиса (1977).
Применительно к древесным породам нами предпринята попытка использования данных методов на ЭВМ по программе, разработанной автором совместно с В. Т. Рыжковой, и был проведен сравнительный анализ насаждений, различных по своему происхождению (Егоров М. Н., 1986, 1988, 1989, 1993 и 1995). Для классификации объектов (для нашего случая деревьев на пробной площади) использовался Q-метод кластерного анализа и коэффициент сходства cos Q. В качестве объекта исследований была взята сосна обыкновенная из Среднеуральской и Центрально-Черноземной популяций, где и были заложены одновозрастные пробные плошади в уже сформировавшихся природных насаждениях и одновозрастных с ними культурах сосны закладки 1861 и 1864 гг. со снятием комплекса показателей их роста и развития.
Итоги анализа выглядят следующим образом: для условий Центрального Черноземья (рис. 4.3 и 4.4) у природного древостоя Х-ось уровня подобия варьирует от -0,2641 до 1,0156; в то время как для культур сосны производства 1861 г. – от -0,2082 до 1,0097. Природный древостой образует 8 кластеров, а культы – 9 кластеров, причем по более высокому значению коэффициента сходства cos Q в культурах деревьев больше, чем в естественном древостое. Из дендрограммы для сосны из Среднего Урала (рис. 4.5 и 4.6) видно, что для природного древостоя Х-ось уровня подобия деревьев варьирует от -0,1456 до 0,9310, тогда как для культур сосны производства 1864 г. – от 0,2121 до 0,9787. Природный древостой образует 9 кластеров, а культуры – лишь 6, причем по более высокому значению коэффициента сходства cos Q (0,9787 против 0,9310) в культурах сосны деревьев значительно больше, чем в естественном сосновом древостое.
Рис. 4.3. Дендрограмма (Х-ось уровня подобия) пробной площади № 11 (Воронежский лесхоз, Углянское лесничество, кв. 46). Естественное сосновое насаждение VII класса возраста

Рис. 4.4. Дендрограмма (Х-ось уровня подобия) пробной площади № 12 (Воронежский лесхоз, Углянское лесничество, кв. 46). Культуры сосны производства 1861 г.

Рис. 4.5. Дендрограмма (Х-ось уровня подобия) пробной площади № 5 (Билимбаевский лесхоз Свердловской области, кв. 52). Естественное сосновое насаждение VII класса возраста
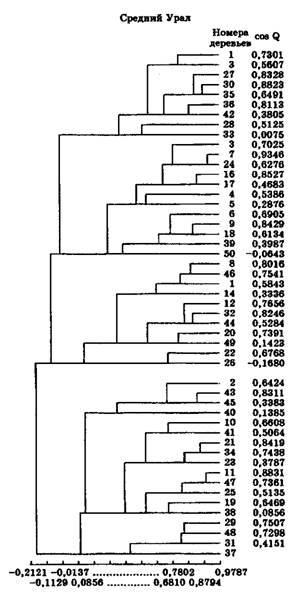
Рис. 4.6. Дендрограмма (Х-ось уровня подобия) пробной площади № 6 (Би- лимбаевский лесхоз Свердловской области, кв. 52). Культуры сосны производства 1864 г.
4.2.2.1. Q-метод кластерного анализа по групповому коэффициенту связи
В основу данного Q-метода кластерного анализа по групповому коэффициенту связи (ГКС) положено заключение о том, что каждый объект рассматривается как отдельная группа. Для n-объектов в начале процедуры будет n групп. Первый шаг классификации заключается в нахождении двух наиболее сходных объектов в матрице значений (n – 1)/2 пар объектов с последующим объединением их в одну группу, после чего остается n – 1 групп. Значение вычислений меры сходства для двух наиболее более сходных объектов рассматривается как характеристика уровня первого шага группировки. На втором шаге объединения возможны два варианта: первый – простое объединение двух объектов в одну группу, как и в первом шаге, или присоединение третьего объекта к образованной на первом шаге группе из двух объектов; для второго варианта нужно определить меру расстояния между группами. В качестве такой меры принимается усредненное внутригрупповое расстояние между парами. Если от этой усредненной меры вычислить косинус, то можно получить среднее значение коэффициента корреляции в группе.
Процесс последовательного объединения групп будет продолжаться до тех пор, пока n-объектов не будут объединены в одну группу. Для n-объектов эта процедура требует n – 1 шагов последовательного объединения. Программа Q-метода кластерного анализа для данного случая составлена на алгоритмическом языке "ФОРТРАН" с использованием транслятора Ф-20. Итогом работы программы является таблица, включающая номер цикла, ГКС – как косинус минимального расстояния уровня шага объединения и порядковые номера элементов объединенных групп.
Данным методом изучены природные и искусственные насаждения сосны в плане их сопоставительной оценки в завершающем возрастном интервале их формирования, роста и развития. По ГКС искусственные насаждения разбились по группам по мере "затухания" сходства следующим образом: по ГКС 0,9995 – 6,5 %, 0,9990 – 0,9995 – 35,5 %, 0,9980 – 0,9990 – 40,5 % и 0,9980 – 17,5 %. У природного же древостоя эти соотношения составили 0; 19,7; 32,8 и 47,5 %. Интервал варьирования ГКС для культур сосны равен 0,9903 – 0,9997, для естественных насаждений – 0,9875 – 0,9994. Количество особей, наиболее сходных по комплексу признаков, в культурах сосны при ГКС 0,9990 составило 42 %, в то время как в природном древостое – 19,7 %, что меньше по сравнению с первыми более чем в два раза. Доля особей с ГКС 0,9980 в культурах сосны – 17,5 %, в естественных же насаждениях – 47,5 %, что указывает на более высокую гетерогенность природных древостоев сосны. Следует отметить, что в природных насаждениях не выявлены группы деревьев с ГКС 0,9995, в то время как в культурах они представлены 6,5 %, что свидетельствует о наличии в них представительства групп особей с более высокой мерой сходства по сравнению с природными древостоями (в анализе использованы данные пробных площадей, заложенных в 130-летнем природном древостое и культурах сосны производства 1864 г.).
Таким образом, анализ сходств (различий) по ГКС показал, что в культурах сосны доля особей с ГКС 0,9980 почти в три раза меньше, чем в природных древостоях, что опять-таки подтверждает мысль о более высокой гетерогенности природных популяций в сравнении с культурами того же возраста.
Проведенные исследования выявили достаточно высокую эффективность данных методов в сравнительном анализе изучаемых популяций, и они вполне могут быть применены ко всем лесным древесным растениям.
4.2.3. Изотропный метод сравнительного многомерного анализа
В последнее время среди специалистов из различных отраслей науки чрезвычайно возрос интерес к применению в своих исследованиях эвристических методов (Плюта В., 1989). Применительно к древесным растениям предпринята попытка использования данных методов и нами. Принимая числовые данные как объект использования сравнительного многомерного анализа (СМА), можно дать информацию об их однородности, определенных локальных сгущениях и разреженностях в изучении совокупности точек-объектов (для нашего случая – деревьев по комплексу структурных признаков). Такую информацию позволяет осуществить анализ следа распределения (Hellwig Z., 1965) или, иначе, – информацию об определенных особенностях распределения многомерной случайной величины.
При определении следа распределения многомерных случайных величин все чаще находят применение методы СМА (Беккер А. В. и др., 1975). Особый интерес к методам СМА вызван тем, что ими можно анализировать расположение точек числовых данных в многомерном пространстве, рассматриваемых как объекты таксономического исследования. Осуществив предварительные операции (выделив набор свойств, характеризующих изучаемые явления, избавившись от единиц измерения свойств и вычислив матрицу расстояний между объектами), приступаем к разделению совокупности объектов на однородные подмножества, используя метод шаров (Bukietynski W., Hellwig Z., Rrolik U., Smoluk A., 1969). Алгоритмическая схема данного метода положена в основу программы для ЭВМ применительно к изучаемой нами совокупности деревьев по комплексу структурных признаков. Для графического изображения нами избрана бинарная индексированная классификация, где каждый узел имеет двух непосредственных преемников (Жамбю М., 1988).
На основе алгоритмической схемы данного метода и разработанной программы были проанализированы практически во всех Возрастных интервалах сосняки естественного и искусственного Происхождений из двух контрастных по условиям местопроизрастания зон (Средний Урал и Центральное Черноземье), позволившие сделать вполне определенные выводы. В качестве примера приведем данные для 130-летних одновозрастных сформировавшихся природных сосняков и культур сосны обыкновенной производства 1864 г. по соотношению выделенных кластеров, составившие 8:16 (т.е. 1:2) при р = 1,13 и 1,16 для условий Среднего Урала. В Центральном Черноземье для 130-летних природных сосняков и культур сосны производства 1861 г. соотношение кластеров составило 7:13 при р = 1,36 и 1,19 (р – величина радиуса шара).
В стадии уже спелых насаждений как в природных популяциях, так и в культурах сосны наибольшей информационной ценностью обладают такие диагностические признаки, как объем ствола, высота, диаметр, архитектоника кроны и ее ширина; толщина, строение и цвет коры.
Для иллюстрации можно предложить дендрограммы данных пробных площадей (рис. 4.7 и 4.8), заложенных в природном спелом сосновом древостое и одновозрастных с ним культурах сосны производства 1861 г. в Центральном Черноземье.
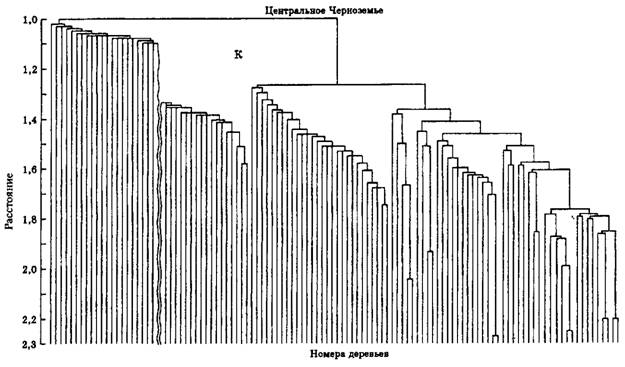
Рис. 4.7. Дендрограмма данных пробной площади № 12, заложенной в культурах сосны производства 1861 г. (Воронежский лесхоз Воронежской области, кв. 46). Первое подмножество-кластер приведено в "разорванном" виде из-за его значительного размера по горизонтали
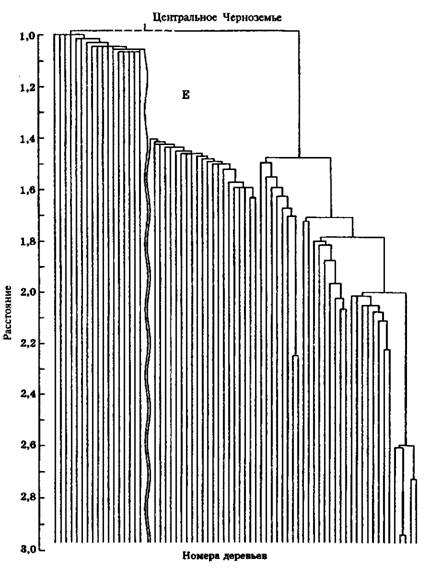
Рис. 4.8. Дендрограмма данных пробной площади № 11, заложенной в Природном спелом сосновом древостое (Воронежский лесхоз Воронежской области, кв. 46). Первое подмножество-кластер приведено в "разорванном" виде" из-за его значительного размера по горизонтали
Рассмотренный изотропный метод СМА показал весьма большую разрешающую способность в изучении фенотипической структуры сравниваемых популяций сосны, и он с успехом может быть применен и к другим древесным породам. Таким образом, независимо от лесорастительных зон (Средний Урал и Центральное Черноземье) и происхождения сосняков (природные насаждения и культуры) можно утверждать, что процесс их дифференциации и образования окончательно устоявшихся кластеров практически идентичен.
4.2.4. Координатный метод комплексной оценки
Данное исследование заключалось в разработке метода и на его основе – математической модели для комплексной оценки лесных древесных пород (Булыгин Ю. Е., 1978, 1992). В основу метода положена известная из курса аналитической геометрии теорема о расстояниях между двумя точками, заданными своими координатами относительно декартовой прямоугольной системы координат в трехмерном пространстве, равных квадратному корню из суммы квадратов разностей соответствующих координат этих точек (Корн Г. и Корн Т., 1984). В качестве координат точек многомерного Евклидова пространства Еn в лесоводственном смысле понимается совокупность показателей признаков и свойств исследуемых древесных растений, которые, в свою очередь, с позиций аналитической геометрии и векторной алгебры можно рассматривать как точки или векторы, заданные своими координатами в пространстве.
Математическая модель в данном случае имеет вид
где Pi – расстояние от точки, обозначающей i-ю форму, до точки или формы-эталона: аi1, ai2,..., аin (i = 1, 2, 3,..., n) – набор координат i-й формы в пространстве Еn. Метод был реализован в соответствии с данной алгоритмической схемой нахождения ?i. Распределение селекционных категорий сосны у сформировавшегося по продуктивности спелого природного древостоя по комплексу структурных признаков с учетом оценки pj приведено в табл. 4.1. Этот показатель позволил всю выборку деревьев разделить на три категории: лучшие, средние и худшие. К примеру, к категории лучших отнесена сосна с толстой и средней толщины корой с густой и средней густоты кроной с пластинчатой (пластинчато-чешуйчатой) бледно-коричневой корой, и ей следует отдать предпочтение в селекционном отношении при создании будущих высокопродуктивных насаждений сосны и внедрении в производство.
Таблица 4.1
Распределение селекционных категорий сосны по продуктивности по ряду структурных признаков с учетом оценки р, (Свердловская область, Билимбаевский лесхоз, Первоуральское лесничество, кв. 52, пробная площадь М 5, возраст – 130 лет)


Если проанализировать продуктивность сосны по отдельным структурным признакам, то по толщине коры в категорию лучших попадают формы сосны с толстой и средней толщины корой, в категорию средних – с тонкой корой. По плотности кроны к категории лучших относятся сосны средней густоты и густой кроной, к худшим – с редкой кроной; по архитектонике кроны к лучшим относятся сосны с яйцевидной и шатровидной кронами, к средним – с конусовидной и цилиндрической кронами; к худшим – с пирамидальной и флаговидной кронами; по строению грубой корки к лучшим отнесены сосны с пластинчатой, пластинчато-чешуйчатой корой, к средним – с чешуйчато-пластинчатой и к худшим – с чешуйчатой корой; по цвету коры к лучшим отнесены деревья с бледно-коричневой корой, к средним – с коричневой корой и к худшим – все остальные особи из имеющейся и анализируемой цветовой гаммы с светло-красной, бледно-серожелтой, коричнево-желтой, серой, желтой и светло-желтой окраской.
Выявленные перспективные категории сосны обыкновенной могут быть использованы в практике лесохозяйственного производства для испытания их по семенному и вегетативному потомству, а также для внедрения лучших хозяйственно ценных форм при создании искусственных насаждений в будущем.
4.2.5. Показатели популяционной изменчивости по полиморфным признакам
Как отмечал Л. А. Животовский (1984, с. 7), "в современных популяционных работах особей исследуют по большому числу разнообразных признаков. Это позволяет проводить детальный анализ структуры популяций, оценивать значимость микроэволюционных факторов, прогнозировать генетические изменения в популяциях... Это преимущественно моногенные признаки и признаки, определяемые блоком тесно сцепленных генов. Они характеризуются дискретным проявлением, и особи однозначно классифицируются по ним четко на различные генотипические классы".
Сравнительный фенотипический анализ сформировавшихся природных древостоев и одновозрастных с ними культур сосны обыкновенной производства 1861 и 1864 гг. из Центрально-Черноземной и Среднеуральской популяций на основе показателей по полиморфным признакам Л. А. Животовского (1979, 1980, 1982) дан в табл. 4.2.
Таблица 4.2
Показатели сравнительного внутрипопуляционного разнообразия природных сосняков и культур по среднему числу и доле редких морф по совокупности маркерных признаков (Центральное Черноземье и Средний Урал)


Для наглядности приведем значения показателей популяционной изменчивости (?, h, r, I) для уже сформировавшихся и прошедших все возрастные стадии роста, развития и становления спелых сосняков, различных по происхождению и зонам произрастания, указывающих на достоверность различий между ними. В частности, при сравнении культур 1864 г. и природных древостоев из Среднеуральской популяции по ? всего комплекса признаков (толщина, строение и цвет коры, очищаемость ствола от сучьев, плотность и архитектоника кроны) ? = 24,532 ± 0,652 и 21,215 ± 0,788 при tf = 3,24* и h = 0,124 ± 0,023 и 0,078 ± 0,034 при tf = 1,12; у Центрально-Черноземной популяции для культур 1861 г. и природных древостоев ? = 13,293 ± 0,356 и 14,653 ± 0,495 при tf. = 2,23* и h = 0,114 ± 0,023 и 0,186 ± 0,027 при tf = 2,12*.
Если проанализировать данные по совокупности независимых признаков, то по показателю сходства популяций r для Среднеуральской популяции имеем значения r = 0,896 ± 0,0116 и критерий идентичности I = 190,0*; для Центрально-Черноземной популяции r = 0,928 ± 0,0076 и I = 229,6*. Если провести сравнение природных популяций и культур сосны из таежной и лесостепной зон, то получим следующие данные: для природных сосняков ? = 20,858 ± 0,657 и 20,385 ± 0,516 при tf = 0,57 и h = 0,166 ± 0,026 и 0,114 ± 0,023 при tf = 1,49; для культур сосны 1864 и 1861 гг. ? = 24,532 + 0,652 и 13,293 ± 0,356 при tf =15,13* и h = 0,124 ± 0,023 и 0,114 ± 0,023 при tf = 0,31. Для природных сосняков r = 0,922 ± 0,0057 и I = 376,0*; для культур – r = 0,855 ± 0,032 и I = 284,0*.
Следовательно, во всех рассмотренных вариантах достоверность различий между ними, оцениваемая по I показателю r, очевидна (* – различия достоверны). (Показатели Л. А. Животовского: ? – среднее число морф; h – доля редких морф; r – показатель сходства популяций; I – критерий идентичности.)
Следует отметить, что независимо от рассматриваемых контрастных лесорастительных зон отчетливо выделяются вариации сосны по строению, толщине и цвету коры, плотности, щирине и архитектонике кроны, очищаемости ствола от сучьев и при сравнении популяций сосны в связи с ее происхождением по данным признакам выявлены следующие особенности:
– по толщине коры по значению ? и h различия достоверны для спелых сформировавшихся сосняков и культур сосны из таежной (Средний Урал) и лесостепной (Центральное Черноземье) зон;
– по строению коры для спелых природных сосняков и культур сосны из данных зон как по ?, так и по h различия достоверны;
– по очищаемости ствола от сучьев для природных сосняков из указанных зон как по ?, так и по h различия недостоверны;
– по плотности кроны различий между сосняками этих зон нет;
– по архитектонике кроны у природных сосняков и культур обеих зон различия достоверны по значению ?, по h их нет:
– по цвету коры нет достоверных различий по ? между природными сосняками из таежной и лесостепной зон, по значению h различия достоверны, и лишь для культур сосны 1864 и 1861 гг. различия достоверны по ?, но недостоверны по h.
Если провести сравнение природных и искусственных сосняков по целой группе независимых признаков среднего числа морф ? и доле редких морф h, то различия достоверны для спелых сформировавшихся природных сосняков из рассматриваемых зон. По показателю же сходства популяций г и критерию идентичности различия I между природными сосняками Среднего Урала и Центрального Черноземья на 1 %-ном уровне значимости достоверны по толщине коры; по строению коры – на 0,1 %-ном уровне значимости; по очищаемости ствола от сучьев достоверных различий нет; по плотности кроны различия достоверны на 1 %-ном уровне значимости; по архитектонике кроны различия также достоверны; по цвету коры различия налицо на очень высоком пороге доверительной вероятности. По значению I на 0,1 %-ном уровне значимости сопоставляемые сосняки достоверно отличаются друг от друга.
Следовательно, по степени сходств и различий сравниваемые сосняки из лесной и лесостепной зон имеют различия между собою. Предложенный метод сравнительного анализа популяций по комплексу полиморфных признаков является также одним из универсальных и эффективных подходов к изучению структуры популяций древесных растений. Он позволяет быстро и результативно находить нужные исследователю решения.
4.3. ОСОБЕННОСТИ ФЕНОТИПИЧЕСКОЙ СТРУКТУРЫ СОСНЯКОВ В СВЯЗИ с ДИССИММЕТРИЧЕСКОЙ ИЗМЕНЧИВОСТЬЮ ГЕНЕРАТИВНЫХ ОРГАНОВ
Диссимметрия дословно трактуется как нарушение, расстройство симметрии. Как отмечал И. В. Гете, "все формы похожи, и ни одна не одинакова с другой: так весь хор указывает на тайный закон" (Канаев И. И., 1964, с. 85). Хотя левизна-правизна филотаксиса была открыта Шарлем Бонне в 1754 г., однако лишь в последние 30 лет началось систематическое изучение данного явления. В. И. Вернадский (1935, с. 23) писал, что "научная теоретическая мысль только подходит к области явлений правизны и левизны. Это почти непочатое поле процессов, которое мне представляется затрагивающим основные проблемы естествознания" и что "диссимметрия – левизна-правизна – одно из основных проявлений жизни" (1967, с. 106).
На явление левизны-правизны обратили внимание также многие исследователи: Д. В. Наливкин (1925), Н. П. Кренке (1933), Г. В. Вульф (1952), Ю.А. Урманцев (1961), И. И. Шафрановский (1985) и др. Высказывалась даже мысль, что большую роль в левом и правом развитии у растений играют однонаправленность вращения земного шара, условия освещенности растения солнцем, географические влияния и ряд других причин.
Как справедливо отмечает А. В. Хохрин (1977, с. 1), "совершенно неизученной оставалась диссимметрическая форма изменчивости древесных растений, связанная с явлениями левизны-правизны. Актуальность данной проблемы давно назрела".
За последние два десятилетия XX в. перечень опубликованных работ по древесным породам говорит о том, что интерес к этим исследованиям не только угас, но и наоборот, они развертываются все полнее и шире (Бакшаева В. И., 1971; Ившин В. Н. и Хо)фин А В., 1973; Голиков А. И., 1979; Голикова В. В. и Голиков А. М., 1981; Хохрин А В., 1981; Захаренко Г. С., 1986; Толстопятенко А. И., 1989, 1991; Голиков А. М. и Карцев А. Д. и др., 1992; Егоров М. Н., 1993; Жестерев А М., 2000 и др.). Из этих работ видно, насколько актуальна разработка данного раздела ботаники и дендрологии для познания функциональной роли в жизни древесных растений левого и правого филотаксиса (зеркальной изомерии), проявляющегося на морфологическом уровне в строении побегов и шишек, или, по А. В. Хохрину (1977, с. 4), "внутривидовая диссимметрическая изменчивость древесных растений исследуется впервые, поэтому... развивает новое теоретическое направление – стереобиологию древесных растений".
Однако работ в плане сопоставительного анализа по диссимметрической изменчивости популяций древесных растений в зависимости от их происхождения нами не выявлено; поэтому разработка этого направления исследований представляется крайне необходимой для познания микроэволюционных процессов, складывающихся в природных и искусственных популяциях. Ближайшую цель исследования мы видим в выявлении функциональной роли в жизни древесных растений левого и правого филотаксиса, проявляющегося на фенотипическом уровне в строении шишек из различных популяций. В связи с этим были поставлены следующие конкретные задачи:
– исследовать диссимметрическую изменчивость сосны обыкновенной в зависимости от ее происхождения по левизне-правизне шишек;
– изучить закономерности распределения левых и правых форм Шишек в различных популяциях и экологических условиях, степень их сходства и различия по рассматриваемым признакам;
– исследовать рост энантиоморф сосны левых и правых форм для выявления преимуществ той или иной формы;
– проанализировать различия левых и правых форм сосны в сопоставлении с другими хвойными породами в природных древостоях и культурах по структурным признакам зрелых шишек.
Конечная цель исследования заключается в установлении неизвестных ранее закономерностей в биологии сосны и структуре ее популяций по диссимметрической изменчивости шишек в зависимости от происхождения, чтобы конкретизировать существующие представления о внутривидовой изменчивости и выяснить преимущество тех или иных форм для их внедрения в лeсохозяйственную практику. Из всего имеющегося наличия проявлений левизны-правизны у сосны был выбран один признак – диссимметрия филотаксиса шишек.
Наряду с сосной обыкновенной (Pinus sylvestris L.) в опытные работы были включены сосна крымская (Pinus pallasiana Lamb.), сосна Станкевича (Pinus Stankewiezi Suk.), лиственница Сукачева (Lariks Sukaczewii Djil), ель сибирская (Picea obovata Ldb.). Объекты исследования представлены насаждениями всего возрастного диапазона от I класса возраста до спелых и перестойных, различных по происхождению, экологическим и лесорастительным условиям местопроизрастания: лесная и лесостепная зоны – Урал, европейская и азиатская части России (Свердловская, Пермская, Тюменская, Воронежская и Кировская области) и Крым. Всего изучено 883 выборки из различных популяций хвойных пород. Диссимметрическая изменчивость изучалась с учетом эндогенной, индивидуальной и географической форм изменчивости (Мамаев С. А., 1972) по методике А. В. Хохрина (1970).
При изучении энантиоморфизма сосны обыкновенной в различных популяциях из сравнения сопоставляющих выборок по процентному соотношению правых (D) и левых (L) форм шишек видно, что достоверность различий для природных популяций очевидна по критерию t-Стьюдента для Р = 0,999 в выборках, приуроченных к экстремальным условиям местопроизрастания, – сосняк по болоту и гранитным обнажениям скал (табл. 4.3). Для сосны крымской достоверность различий по критерию t-Стьюдента при Р = 0,95 доказывается для вариантов с L- и D- формами шишек в естественных сосняках и культурах (процентное соотношение L- и D- форм в естественных древостоях и культурах составило соответственно 65,9 и 34,1 %; 60,6 и 39,4 %).
Таблица 4.3 Соотношение энантиоморф сосны обыкновенной по правизне-левизне шишек для различных ее популяций

В зависимости от формы апофизов на концах семенных чешуй шишек отчетливо подмечаются три категории сосен: с гладкими, бугорчатыми и крючковатыми шишками, хотя имеют место и переходные формы (Егоров М. Н., 1976). Чтобы выяснить, не накладывают ли отпечаток те или иные формы сосны по данному структурному признаку на соотношение правых и левых форм, был взят исходный материал как в природных насаждениях и культурах, так и в привитых архивно-маточных плантациях из Свердловской, Кировской и Воронежской областей. Оказалось, что при анализе культур сосны достоверность различий приведенных выборок по критерию t-Стьюдента очевидна лишь для гладкой формы апофиза и для правой формы гладкого и бугорчатого апофиза; для остальных вариантов при Р = 0,95 она не подтверждается. У природных сосняков достоверность различий для всех выборок не доказана, так как tf &;lt; tst Для выяснения складывающихся особенностей в соотношении
L- и D-форм при вегетативном потомстве были заложены архивно-маточные плантации в Сомовском лесхозе Воронежской области от деревьев-маточников из Вятско-Полянского лесхоза Кировской области и Билимбаевского лесхоза Свердловской области. Окраска шишек варьирует с богатой гаммой цветов и оттенков, и отсюда вполне правомочно возникает вопрос о степени влияния цвета шишек на распределение левых и правых форм. Оказалось, что по форме апофиза шишек различия достоверны лишь в одном случае для бугорчатых шишек из Вятско-Полянского лесхоза по критерию t-Стьюдента при Р = 0,95; по цвету же шишек достоверность различий не выявлена во всех рассмотренных случаях. Для сравнения аналогичные исследования были проведены с елью сибирской из различных условий ее местопроизрастания (Пермская область – Пермский и Соликамский лесхозы; Свердловская область – Староуткинский и Билимбаевский лесхозы), где достоверных различий не обнаружено. В итоге получается, что выявленные закономерности в соотношениях L- и D-форм шишек по их левизне-правизне присущи не только для сосны обыкновенной, но и для других хвойных пород независимо от мест их произрастания.
Рассмотренный экспериментальный материал позволил сделать следующие выводы:
– из исследованных семи природных популяций сосны обыкновенной, резко различающихся по своим природно-климатическим условиям (Крым, Средний Урал, Предуралье, Центральное Черноземье), достоверность различий энантиоморф сосны по левизне-правизне шишек и их весовым соотношениям между собою не выявлена, за исключением выборок, взятых в экстремальных условиях местопроизрастания (Средний Урал – сосняк по болоту и фанитным обнажениям скал), где соотношение L- и D- энантиоморф составило 69,5:30,5 и 16,5:83,5 %;
– при сравнении культур сосны обыкновенной в лесной и лесостепной зонах (Средний Урал – культуры 1864 г. и Центральное Черноземье – культуры 1861 г.) энантиоморфизм сосны по L- и D-формам шишек существенных достоверных различий не выявил – примерное соотношение L- и D- форм тяготеет к пропорции 0,5:0,5. Следовательно, независимо от различия лесорастительных зон и наложения антропогенного фактора заметного эволюционного сдвига по рассматриваемым признакам не произошло;
– при сопоставлении одновозрастных насаждений, различных по происхождению, в пределах одного района их местопроизрастания достоверных различий L- и D-форм по их процентному соотношению не выявлено, и оно практически для всех случаев близко к 0,5:0,5;
– несколько иное соотношение получено при сравнении L- и D-форм у сосны крымской, где в природных сосняках оно составило 65,8:34,2 %, т.е. почти 2:1; в культурах – 60,6:39,4%, или 1,5:1. Достоверность различий по критерию t-Стьюдента при Р = 0,95 очевидна для L- и D-форм в культурах и D-формы культур и L-формы природных сосняков. Здесь явно просматривается преобладание L-форм над D-формами в 1,5 – 2,0 раза. Для условий изолированной популяции сосны крымской в ее специфических природно-климатических условиях, очевидно, произошел некоторый сдвиг по разбалансированию равновесия L- и D-форм;
– при изучении L- и D-форм в культурфитоценозах в условиях Центрального Черноземья по строению апофиза семенных чешуй достоверных различий трех категорий шишек (плоские, бугорчатые и крючковатые) по критерию t-Стьюдента не выявлено, и соотношение их тяготеет к пропорции 0,5:0,5 (43,4:56,6 % для гладкого апофиза; 51,8:48,2 % – бугорчатого и 50,4:49,6 % – крючковатого);
– при сравнении L- и D-форм в природных древостоях в условиях лесостепи (Центральное Черноземье) по апофизу семенных чешуй достоверность различий рассмотренньк трех категорий шишек не установлена, и их соотношение близко к пропорции 0,5:0,5 (гладкие – 51,9:48,1 %; бугорчатые – 45,3:54,7 %; крючковатые – 53,0:47,0 %);
наблюдается смещение в сторону диспропорции L- и D-форм по строению апофиза в прививках архивно-маточной плантации культур сосны обыкновенной из Среднего Урала и природных насаждений из Предуралья, что вполне может быть объяснено тем, что на плантацию попали деревья с вполне определенной структурой по L- и D-формам, и они, в свою очередь, наложили свой отпечаток на соотношение данных форм в прививках и на семенное потомство;
– при анализе природных древостоев сосны по строению апофиза шишек из Центрального Черноземья соотношение L- и D-форм у гладких, бугорчатых и крючковатых форм шишек Достоверность различий выявлена лишь в двух случаях – между L- и D-формами гладких шишек и D-формами у гладкой и бугорчатой категорий шишек. Соотношение L- и D-форм для гладкой категории составило 0,52:0,48; бугорчатой – 0,45:0,55; крючковатой – 0,53:0,47;