Памяти учителя, видного ученого
в области лесоведения и лесоводства, профессора Нила Алексеевича Коновалова посвящается
ПРЕДИСЛОВИЕ
Монография посвящается одному из новых оригинальных направлений и исследований лесных популяций древесных растений – фенетике древесных пород. Становление фенетики как нового направления генетических исследований в нашей стране связано с "Законом гомологических рядов в наследственной изменчивости" (Вавилов Н. И., 1920), в котором фактически дана формулировка теоретической основы фенетики популяции растений. Н. И. Вавилов первым осознал необходимость получения точных данных по видам и популяциям, генетическое изучение которых не проводилось или сопряжено с определенными трудностями в методологическом плане.
Известно, что в настоящее время доля видов, изученная генетически, ничтожно мала – не более 0,02 % общего количества существующих видов и соответственно популяционно-генетически исследованных – около 0,007 %. В силу сложностей изучение больщинства видов на генетическом уровне (куда, в частности, отнесены и древесные растения) становится весьма проблематичным. Генетический "вакуум", образовавшийся у древесных пород ввиду длительной смены у них поколений, не позволяющий использовать традиционные методы генетики и селекции (направленные скрещивания и анализ в следующих поколениях) для улучшения хозяйственно ценных признаков деревьев, и неотложность селекционной работы на достаточно надежной генетической основе вынуждают изыскивать новые подходы к реализации программ селекции.
Предмет, цели и методы фенетики популяций как направления исследований, развивавшегося на стыке генетики и классической зоологии и ботаники, впервые были сформулированы Н. В. Тимофеевым-Ресовским (1973). До ее применения о структуре популяций судили в основном по характеру распределения особей в пространстве. Фенетика же с использованием признаков-маркеров генотипического состава позволяет выявить структуру популяций с неизмеримо большей точностью и наименьшими затратами усилий, давая представление о том, на какие фенетические группировки делится вся совокупность особей. При анализе популяций и внутрипопуляционных группировок используется основное свойство фена (его наличие или отсутствие и частота встречаемости) и именно в широте охвата фенами генотипа одно из определяющих преимуществ фенетического подхода к изучению генотипической структуры популяций по анализу аллелофонда. Структуру монографии определил в общем сам фенетический подход, в котором последовательно рассматриваются узловые вопросы ее содержания.
В "фундамент" работы были положены основополагающие идеи, сформулированные в свое время Н. И. Вавиловым, Н. В. Тимофеевым-Ресовским, А. В. Яблоковым, Н. В. Глотовым, С. С. Шварцем, М. М. Магомедмирзаевым, Л. А. Животовским и другими исследователями. В свою очередь, мы полагаем, что при изучении внутривидовой изменчивости и особенностей фенотипической структуры насаждений и популяций древесных пород должен быть положен именно фенетический подход как наиболее эффективный и перспективный с подключением к нему целого ряда методических решений, основанных на использовании комплекса качественных и количественных признаков структурной, функциональной и биохимической природы их происхождения.
Настоящее издание является основанием для учреждения специального курса по фенетике популяций древесных растений.
Работа выполнена в лаборатории лесной селекции ФГУП "Научно-исследовательский институт лесной генетики и селекции" Министерства природных ресурсов Российской Федерации. Автор выражает надежду, что представленная широкому кругу читателей монография окажется полезной, и будет благодарен за аргументированные и конструктивные замечания и предложения по ее дальнейшему совершенствованию.
Не исключено, что именно фенетика популяций окажется в ближайшем будущем лидирующим направлением исследований в популяционной биологии.
А. В. Яблоков (1983)
ВВЕДЕНИЕ
Жизнь на планете Земля характеризуется прежде всего дискретностью и представлена особями, как правило, разбитыми на отдельные группы, образующие систему определенных совокупностей-таксонов; причем основные уровни организации включают молекулярно-генетический, онтогенетический, популяционно-эволюционный, биогеоценотический и биосферный и тесно взаимосвязаны.
Учение о популяции зиждется на трех основных постулатах: динамике численности вида и флуктуации занятой им территории, теории систематики и таксономической практики, микроэволюционных процессах и пусковых механизмах эволюции (Тимофеев-Ресовский Н. В. и др., 1973).
Несмотря на определенные успехи в изучении популяций, от современного толкования эволюционных процессов и факторов на микроэвалюционном уровне до перехода к управляемой эволюции – дистанция огромного размера, и больщинство видов в природе в обозримом будущем не будут изучены на генетическом уровне. В этом понимании единственной "спасательной шлюпкой" может быть лишь фенетический подход, позволяющий проводить изучение популяций, опираясь на экспериментальные данные морфологических, экологических и этологических исследований на неограниченном спектре видов растительного и животного мира; именно фенетике в популяционной биологии в ближайшем будущем будет отведено место лидера как самостоятельного направления исследований (Яблоков А. В., 1978).
Организация жизни на нашей планете такова, что генотип особи вида реализует ее фенотип в практически неограниченном спектре внешних условий и зачастую идентичные фенотипы являются "продуктом деятельности" различных генотипов, и наоборот, различные фенотипы могут быть сформированы на достаточно близкой генетической основе. Однако при этом следует отметить существенное различие между понятиями фенетики как генетико-эволюционного направления в изучении популяций и фенетики как раздела теоретической систематики.
Разрозненный и фрагментарный отрывочный материал по фенетике древесных растений, начиная с 70-х гг. XX в. и по настоящее время, к сожалению, не дает цельного и объемного представления об этом перспективном и многообещающем направлении исследований, и данная сводка может рассматриваться как одна из первых попыток саккумулировать весь накопившийся круг сведений и занять ей свое место в ранге автономного самостоятельного направления в биологической науке вообще и применительно к лесным биогеоценозам, в частности.
Актуальность изучения фенофондов и фенотипической структуры природных популяций и культур древесных растений, состояния и перспектив развития исследований в данном направлении продиктована как чисто научными, так и, в гораздо большей степени, производственными запросами интенсивно развивающегося лесного хозяйства страны. В частности, практическое применение итогов исследований нацелено на разрешение задач по научно обоснованному формированию оптимального с селекционной точки зрения качественного состава и продуктивности культур из хозяйственно ценных форм и селекционных категорий, выявленных на основе фенетического подхода.
ГЛАВА 1 ФЕНЕТИКА ПОПУЛЯЦИЙ ДРЕВЕСНЫХ РАСТЕНИЙ
1.1. МЕСТО И РОЛЬ ПОПУЛЯЦИИ в эволюции
Современный этап в развитии учения об эволюции характеризуется возникшим в 30-е гг. XX в. изучением микроэволюционных процессов, явившимся результатом взаимопроникновения классического дарвиновского эволюционного учения и генетики (Dobzhansky Th., 1937; Тимофеев-Ресовский Н. В., 1958). Микроэволюция исследует популяционно-генетические процессы в их динамике, ведущие к формированию явления адаптации, закреплению внутрипопуляционных проявлений полиморфизма, началу формо- и видообразования и других пусковых процессов эволюции.
Однако имеющиеся данные о формовом составе главных лесообразующих пород, являющихся предметом изучения в настоящем исследовании, не дают полного представления о структуре их популяций и закономерностях микроэволюционных процессов, и в этом направлении предстоят дальнейшие исследования.
С. А. Мамаев (1974) отмечает, что важнейшая задача, которую решает внутривидовая систематика растений, – это проблема микроэволюции в научно-теоретическом аспекте, а в практическом смысле – научно-методическое обоснование селекции растений. К. М. Завадский и Э. М. Колчинский (1977) под микроэволюцией понимают совокупность преобразований популяций, протекающих в рамках вида, и ее изучают ряд разделов биологии, занимающихся сравнительным анализом популяций и закономерностей внутривидовой изменчивости. В эволюционных исследованиях далее неподразделяемой, элементарной эволюционной единицей является популяция как одна из главных элементарных структур организации жизни на Земле, формулировка понятия которой явилась наиболее крупным эмпирическим обобщением биологии XX в.
Место и роль популяции в эволюции органического мира достаточно отчетливы, и биологи-эволюционисты приходят почти к однозначным мнениям, что она является "элементарной размножающейся единицей вида" (Завадский К. М., 1961), "элементарной единицей вида" (Сукачев В. Н., 1963; Корчагин А. А., 1964), "популяция – первая ступень, где разыгрываются явления эволюции" (Дубинин Н. П., 1966), "популяция – кузница, где естественный отбор творит новые формы" (Лобашов М. Е., 1967), "наименьшей единицей эволюции" (Шмальгаузен И. И., 1968; Шварц С. С., 1969), "элементарной эволюционной структурой.., самой мелкой из реально существующих в природе биохорологических групп особей" (Тимофеев-Ресовский Н. В. и др., 1969).
Л. Ф. Правдин (1974) отмечает, что в имеющихся публикациях внимание было уделено в основном анализу природных популяций насекомых и растений с непродолжительным периодом жизни одного поколения, и лишь за последние годы природные лесные популяции стали объектами детального анализа изменчивости признаков на достаточно обширной территории их ареалов (Sokolowski S., 1931; Драгавцев В. А., 1962, 1963, 1972, 1994; Dragavtsev V. А., 1995; Петров С. А., 1964, 1970, 1975; Правдин Л. Ф., 1964, 1965, 1967; Мамаев С. А., 1968, 1969, 1972; Ирошников А. И., 1964, 1974, 1985; Семериков Л. Ф., 1971, 1975, 1986; Котов М. М. и др., 1977; Махнев А. К., 1982; Царев А. П., 1985; Чернодубов А. И., 1996; Егоров М. И., 1994, 1997, 2001 и др.). Правда, он уточняет, что использование термина "популяция" к лесным культурам, созданным человеком, и где уже был произведен искусственный отбор и возраст которых определяется одним поколением, должно быть ограничено или применяться с соответствующей оговоркой. Успешное развитие теории генетики популяций и новых подходов к сбору и анализу данных по изменчивости древесных пород позволило уже сейчас сделать некоторые обобщения, полезные для понимания эволюции видов и выработки программы и методов исследований популяций древесных растений (Ирошников А И., 1970, 1974; Правдин Л. Ф., 1964, 1967; Мамаев С. А., 1965, 1969; Петров С. А., 1964, 1974 и др.).
Таким образом, изучение изменчивости внутри вида на уровне популяции является новым и перспективным направлением научных исследований на базе уже имеющихся работ по формовому разнообразию древесных растений и познанию закономерностей процессов эволюции вида, а также выявлению его потенциальных возможностей для селекции. Свойства же популяции определяются интефацией процессов и явлений на трех уровнях организации жизни: на клеточно-молекулярном уровне "записана" генетическая информация популяции; на онтогенетическом уровне она реализуется в фенотипах и, наконец, на популяционно-биогеоценотическом уровне она преобразуется (Тимофеев- Ресовский Н. В. и др., 1969).
Разработка региональных программ генетических ресурсов группой экспертов ФАО была предпринята в 1968 г. и продолжена рядом других международных организаций с участием России и стран ближнего зарубежья. Первым важнейшим шагом в разработке этих программ является определение популяционной структуры вида. Учение о популяционной структуре вида базируется на двух направлениях биологии – на внутривидовой систематике, получившей свое развитие после Ч. Дарвина в связи с кризисом типологической концепции вида, и на популяционной и эволюционной генетике, основы которой заложил С.С.Четвериков (1926). Для познания генетической структуры популяции непреходящее значение имеют работы W. Weinberg (1908) и G. Н. Hardy (1908).
В изучении генетической структуры популяций в течение более чем полувекового развития экспериментальной популяционной генетики были достигнуты большие успехи и выявлена огромная генетическая гетерогенность природных популяций (Dobzhansky Th., 1937, 1970; Дубинин И. П., 1966; Меттлер Л. и Грег Г., 1972; Тимофеев-Ресовский Н. В. и др., 1958, 1973 и др.).
Следует подчеркнуть, что изучение популяционной структуры вида является одной из ключевых проблем в лесоведении, лесной фенетике, генетике и селекции, и актуальность ее определена, в первую очередь, запросами практической селекции и генетического ресурсоведения, потребностями ведения высокопродуктивного лесного хозяйства, лесного семеноводства и лесосеменного районирования.
1.2. ИСТОРИЯ РАЗВИТИЯ ИССЛЕДОВАНИЙ
Исследования растений по дискретным признакам уходят своими корнями ко второй половине XVIII в., в работы ботаников И. Г. Кельрейтера, О. Сажре и Т. Найта, которые во многом предопределили открытие закономерностей наследования Г. Менделем (1865). К необходимости учета "единичных признаков", "единиц характеристики" пришел и Г. Ф. Осборн. Н. И. Вавилов первым осознал необходимость получения данных по видам и популяциям, генетическое изучение которых затруднено или при имеющихся на данный момент научных подходах не реально. В. Иоганнсен (1909) впервые в генетическую терминологию вводит понятие "фен", а А. С. Серебровский (1927) в популяционных исследованиях выделяет геногеографию в самостоятельное направление. Тем не менее о фенетике как автономном направлении исследований можно говорить лишь начиная со второй половины XX в. с работ Н. В. Тимофеева- Ресовского и А. В. Яблокова (1973).
Исследования непосредственно по фенетике древесных пород носят явно "затяжной" характер (табл. 1.1). Лишь начиная с 1973 г. они заявляют о себе с точки зрения как научной, так и практической значимости. А. И. Ирошников, С. А. Мамаев, Л. Ф. Правдин и М. А. Щербакова (1973) публикуют "Методику изучения внутривидовой изменчивости древесных пород", где при описании структурных, функциональных и качественных признаков они отмечают, что "все признаки, по возможности, характеризуются количественными показателями. В случае относительной дискретности признаков необходима унификация параметров. При непрерывной изменчивости необходима унификация числа классов значения признака" (с. 13), т.е. здесь практически уже вовлечены в научный анализ фены как дискретные признаки.

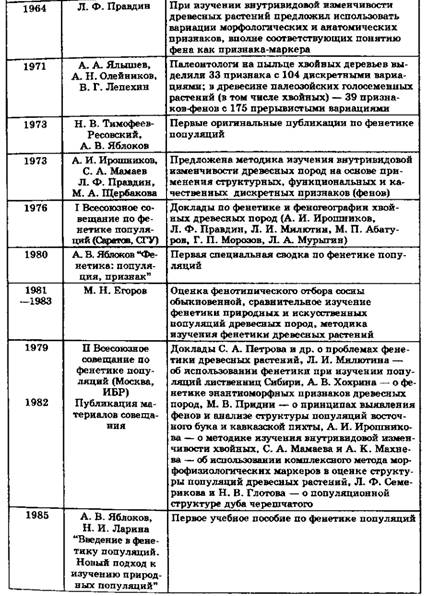

В 1976 г. в Саратове состоялось I Всесоюзное совещание-семинар по фенетике популяций, где, в частности, были впервые представлены работы по фенетике древесных растений – в основном, доклады по фенетике и феногеографии хвойных пород. Интерес к древесным растениям в первую очередь был вызван в связи с практической трудностью изучения их на генетическом уровне в онтогенезе. В материалах II Всесоюзного совещания по фенетике популяций (Москва, 1979) нашли отражение работы, посвященные преимущественно задачам и методам фенетических исследований популяций древесных растений. Достаточно широко представлены работы по фенетике древесных пород на III Всесоюзном совещании (Саратов, 1985), где нашли отражение исследования по фенетике популяций основных лесообразующих пород страны. Обширный перечень работ по фенетике популяций древесных растений был на IV Всесоюзном совещании (Борок, 1990), затрагивающий вопросы как методического плана, так и конкретные результаты изучения фенотипической структуры популяций древесных пород, а также перспективы использования фенетических признаков в качестве маркеров генотипической структуры популяций. Популяционную фенетику естественных и искусственных насаждений сосны обыкновенной анализирует М. Н. Егоров (1997), и им же впервые с применением фенетического подхода к изучению внутривидовой изменчивости и популяционной структуры сосны обыкновенной защищена докторская диссертация на тему: "Фенотипическая структура естественных популяций и культур сосны обыкновенной (на примере сосняков Среднего Урала и ЦЧО)" (1997).
Из приведенного перечня работ видно, что объемы исследований по фенетике популяций древесных растений от года к году непрерывно расширяются; соответственно неуклонно растет научный и практический интерес к ним.
1.3. ОСНОВНЫЕ ПОНЯТИЯ
Фенетика как научное направление – это распространение генетических подходов и принципов на формы, собственно генетическое изучение которых затруднено или невозможно в силу биологических особенностей тех или иных видов.
Предмет фенетики – внутривидовая изменчивость, доводимая в конечном итоге до рассмотрения дискретных альтернативных признаков-фенов.
Методами фенетики являются вычленение в изменчивости изучаемых форм различных фенов, количественное и качественное изучение фенов в популяциях и других группах особей.
Цель фенетики – разработка вопросов микроэволюции, теоретической систематики и ряда других проблем, связанных с популяционным исследованием видов в природе (Тимофеев-Ресовский Н. В. и др., 1973).
Фены – это такие признаки фенотипа, которые характеризуются: а) простой элементарностью, т.е. неразложимостью, неподразделяемостью на составные части без потери качественного своеобразия; б) альтернативностью; в) дискретностью (прерывностью). Последняя характеристика (дискретность), по-видимому, является важнейшей, позволяющей видеть в изучении фенов действенный путь познания дискретного мира живых организмов (Яблоков А. В., 1978).
Внутри фенетики к настоящему времени сформировались три основных раздела: выделение и анализ фенов; изучение фенофондов; феногеография.
1.4. ВНУТРИВИДОВАЯ ИЗМЕНЧИВОСТЬ
Внутривидовая изменчивость в различных формах и вариантах предполагает существование организмов как одномоментное разнообразие генотипов и фенотипов (всех признаков и свойств) у особей и их групп любой степени родства внутри вида и популяции. Древесные растения, как правило, представлены полиморфными видами, имеющими в своем составе несколько четко морфологически отличающихся форм, и полиморфизм вида в широком смысле слова включает в себя все множество форм изменчивости.
Изучение внутривидовой изменчивости древесных растений было начато в конце XIX в. Так, различные формы внутривидовой изменчивости у сосны обыкновенной приводят в своих работах G. Gordon (1880) и F. Schwerin (1906). Обоснование современных представлений о дифференциации вида с применением географо-морфологического, экспериментально-экологического и генетического методов находим у J. Clausen (1922, 1951), J. Clausen, W. Hiesey (1958) и G. Turesson (1922, 1930). О весьма сложном пути от генотипа к фенотипу и взаимодействии генов на клеточном и тканевом уровнях на протяжении онтогенеза, являющимся следствием единства генотипа как "неделимого", указывает С. С. Четвериков (1926). Детальный анализ изменчивости признаков в природных лесных популяциях на достаточно обширной территории их ареалов предпринимает S. Sokolowsky (1931).
Идею комплексного генетико-эколого-геофафического подхода при рассмотрении дифференциации вида предложили М.А.Розанова (1930, 1946) и Е. Н. Синская (1938, 1948, 1963). Е. Ford (1940, 1971) в популяции обнаружил два вида многообразия: сбалансированный полиморфизм, характеризующийся стабильным численным соотношением между формами, поддерживаемыми в популяции из поколения в поколение, и фанзитивный полиморфизм, отмеченный сдвигами численного соотношения форм в популяции в процессе ее эволюционного преобразования.
Исследования по внуфивидовой изменчивости древесных растений и наследуемости признаков потомством приводят Е Andereson (1947) и В. Lindquist (1948), А. С. Яблоков (1949, 1952 1962), С. С. Пятницкий (1954, 1966), К. Rubner (1957), А. В. Альбенский (1959), Е. Rohmeder, Н. Schonbach (1959), А. Gustafsson (1962), М. М. Вересин (1963), Л. Ф. Правдин (1964), С. А. Мамаев (1965 – 1986) и др. Получению в известной мере косвенных характеристик генотипической изменчивости без смены поколений на основе новых методических подходов в генетике количественных признаков и широкому применению математических методов во многом способствовали исследования V.J. Shrighande (1957), В. А. Драгавцева (1963–1994), К. J. Sakai, Н. Mucaide (1967), С. А. Петрова и В. А. Драгавцева (1969), G. Namkoong, А. S. Squilace (1970), Э. X. Гинзбург и В. А. Драгавцева (1970), Л. А. Животовского и Н. В. Глотова (1976), С. П. Мартынова (1976).
К. М. Завадский (1967), анализируя дифференциацию вида, отмечает, что независимо от уровня организации, способа размножения, величины ареала, численности и других особенностей каждый вид включает различные формы, т.е. дифференцирован. Дифференциация может быть различной по разным показателям, но полностью мономорфных видов в природе не существует. В понятие дифференциации вида входят все типы разновидностей: биохимическая, физиологическая, морфологическая, половая, возрастная, сезонная, связанная с фазами жизненного цикла, экологическая, географическая и др.; прерывистая и непрерывная; адаптивная и неадаптивная; модификационная и генетически стабилизированная; проявляющаяся в обычных условиях или же при нарушениях сложившейся системы скрещивания. Однако о том, что генетическая изменчивость в популяциях всегда выступает в своей фенотипической форме, отмечали еще W. Johannsen (1903, 1909) и М. М. Камшилов (1972).
1.5. ИСПОЛЬЗОВАНИЕ ПРИЗНАКОВ И МЕТОДОВ ФЕНЕТИКИ
На необходимость применения методов фенетики для большинства видов, не пригодных для прямого генетического анализа, указывает Н. В. Тимофеев-Ресовский (1973). И. Д. Юркевич и др. (1974) утверждают, что эффективность селекции в лесоводстве будет достигнута за счет использования формового разнообразия и хозяйственно ценных признаков и свойств популяций главнейших древесных пород, отличающихся сильным полиморфизмом и относительно устойчивыми морфолого-биологическими признаками. Л. Ф. Правдин (1974), анализируя изменчивость, отмечает, что индивидуальная изменчивость служит лишь элементарным источником для начала процесса эволюции в группе. Эволюционирует не особь, а популяция. Изучение изменчивости внутри вида на уровне популяции является новым и перспективным направлением исследований, подготовленным предшествующими работами по изучению формового разнообразия древесных пород.
С. А. Петров (1975) констатирует, что на данный момент уже собран достаточно объемный фактический материал по внутривидовой изменчивости древесных растений, служащий одновременно основой для изучения путей и закономерностей их эволюции и имеющий непреходящее значение для практической селекции. Исследование популяционной структуры и эволюции вида, по Р. Левонтину (1978), может быть представлено процессом превращения изменчивости среди особей в пределах свободно скрещивающейся группы в изменчивость групп в пространстве и времени. Л. Ф. Семериков (1981), однако, отмечает, что изучение внутривидовой изменчивости затруднено многообразием ее форм и категорий, а также выбором самих признаков, которые должны достаточно адекватно характеризовать эколого-генетическую архитектонику вида; при этом трудности прямого генетического анализа у большинства лесообразующих видов накладывают серьезные ограничения. Количественные признаки важны с практической точки зрения, поскольку с ними связаны интересы селекционеров.
Для селекции важно накопление информации в популяциях любых признаков, в том числе прямо не связанных с продуктивностью и устойчивостью (история селекции хлебных злаков и создание короткостебельных сортов пшеницы убедительно доказывают важность изучения любых генетически обусловленных признаков); причем развертывание генетической программы в процессе филогенеза обеспечивает в конечном итоге развитие гармонического фенотипа. Путь от гена к признаку или от генотипа к фенотипу оказывается весьма сложным (гены действуют не только на разные стороны фенотипа в определенное время и нужном месте, но и взаимодействуют между собой).
Свою классификацию форм, типов и категорий изменчивости древесных растений предложил С. А. Мамаев (1972); в частности, по типам изменчивости они подразделяются на внутривидовую и внутри организма; по формам изменчивости – на половую, индивидуальную, хронографическую, экологическую, географическую, гибридогенную и эндогенную; по категориям изменчивости – на структурную, функциональную и качественную. Но тем не менее следует согласиться с мнением С. С. Шварца (1963), отметившего, что проявления внутривидовой изменчивости очень многообразны и сложны и "любая попыгка их классификации будет в какой-то степени односторонней и формальной", и подходы к изучению феномена изменчивости можно проводить с разных позиций (с. 418).
Таким образом, несмотря на кажущееся однообразие в строении особей одной популяции, морфологические различия между ними настолько очевидны, что положены в основу современной систематики растений, и ее рабочей единицей чаще всего выступает не одна особь, а их группа или серия. Популяцию оценивают уже по спектру различий в выражении какого-либо диагностического признака или признаков, и тем более изменчивы прочие морфологические особенности древесных растений, которые не были включены в качестве диагностических. Как отмечал Н. В. Тимофеев-Ресовский (1973), наиболее заманчивым направлением исследований в популяционной морфофизиологии является установление степени сходств и различий между разными популяциями и разными группами популяций внутри вида. Если под индивидуальной изменчивостью имеют в виду изменчивость отдельных особей, то под групповой – те же группы изменений, происходящие одновременно у многих особей, живущих в сходной среде.
В исследованиях структуры вида, начиная с 20-х гг. XX в., уже было обращено внимание на выявление и изучение внутривидовых и внутрипопуляционных групп в ранге морфобиологической единицы. Так, В. В. Станчинский (1927) предложил выделить в дифференциальной изменчивости наряду с циклической индивидуальную и групповую (наследственную и ненаследственную) ее формы. Ю. А. Филипченко (1934) также отмечает две категории изменчивости: индивидуальную и групповую. C. C. Robson, О. W. Richards (1936) разрабатывают свою классификацию изменчивости, где выделяют индивидуальную, групповую и видовую формы. Э. Майр (1947) считает, что наиболее важно изучение индивидуальной изменчивости в пределах популяции и групповой у различных популяций в пределах вида; при этом его взгляды о категориях изменчивости очень близки к мнению Ю. А. Филипченко о формах изменчивости. Е. Н. Синская (1963) среди выделенных ею главных категорий изменчивости называет и групповую изменчивость. По А. А. Парамонову (1967), изменения особей составляют индивидуальную изменчивость, изменения видов – исторические изменения. Причем если в первом случае возникают и нарастают различия между особями однородной группы, то во втором они приводят к различиям между видами.
Более детально к анализу групповой изменчивости подходит К. М. Завадский (1967). В частности, из представленной им системы основных внутривидовых единиц он выделяет морфобиологическую группу и биотип. Под морфобиологической группой, или изореагентом, он понимает группу организмов внутри популяции, имеющих одинаковую или различную наследственную основу, выделяемую морфологически и имеющую сходный ритм развития и одинаково реагирующую на условия среды; под биотипом – элементарную единицу генетической структуры популяции в виде группы организмов, имеющих почти идентичный генотип и отличающийся от другой группы в результате хотя бы одной мутации. Каждый вид включает различные формы, т.е. дифференцирован.
Совокупность групп, уже доказавших свое приспособительное значение, формируется под воздействием противоположно направленных процессов индивидуального и группового отбора, ведущих к адаптивной интеграции и радиации групп внутри вида. Ч. Дарвин (1865) разновидностью называл все формы групповых различий внутри вида. Выявление закономерностей формирования внутривидовых группировок входит в задачу внутривидовой систематики при изучении структуры вида у растений (Мамаев С. А., 1974). Л. Ф. Семериков (1981) указывает, что главное внимание при изучении популяционной структуры должно быть направлено на выявление структуры изменчивости признаков и разграничение разных форм изменчивости – особенно на вычленение генетически обусловленной компоненты – индивидуальной (внутрипопуляционной) и групповой (межпопуляционной) изменчивости. Причем вторая категория групповой изменчивости связана с тем, что популяции, в свою очередь, образуют группы, различающиеся комплексом признаков.
К. М. Завадский и Л. Н. Хохина (1983) под "структурой" вида подразумевают лишь охваченные и уже переработанные отбором и имеющие приспособительное значение совокупности внутривидовых групп. И, наконец, при изучении фенофондов сравниваемых популяций А. В. Яблоков и Н. И. Ларина (1985) рамки групповой изменчивости рассматривают в диапазоне разных популяций, разных групп популяций, разных внутрипопуляционных групп и разных групп особей в одной популяции в пространстве и времени. Нам представляется, что последний подход к анализу фупповой изменчивости является наиболее полным и конструктивным в раскрытии ее значения и места в эволюционном процессе.
Таким образом, исторический экскурс в такой природный феномен, как популяция, и пути ее изучения указывают на признание оной в статусе элементарной единицы вида или, по образному выражению генетика М. Е. Лобашова, – кузницы, где естественный отбор творит новые формы. Рассмотренный перечень методических подходов к изучению популяций таких долгожителей, как древесные породы, не всегда позволяет достичь желаемого результата на генетическом уровне, и поэтому здесь неоценимую услугу может оказать фенетика, позволяющая при гораздо меньших затратах усилий и времени получить достоверную информацию как об отдельных особях, так и о популяции в целом.
1.6. МЕТОДИКА ИССЛЕДОВАНИЙ
Особенностью данной работы является попытка одновременной оценки комплекса количественных и качественных структурных признаков на основе различных методологических подходов и решений, так как работ, всесторонне оценивающих ту или иную форму изменчивости с достаточно большим спектром разнообразных признаков у древесных растений, даже принадлежащих к какой-либо одной категории, практически нет. Лишь глубокое познание сущности изменчивости проявляется в исследованиях максимально возможного комплекса признаков и свойств вида.
Изучение изменчивости вида на уровне популяции применительно к древесным растениям проведено в соответствии с ме- годами, предложенными В. А. Драгавцевым (1963–1995), Л. Ф. Правдиным (1964, 1965, 1967), С. А. Петровым (1964, 1970, 1974), С. А. Мамаевым (1965, 1968, 1969), А. И. Ирошниковым и др., (1973), М. М. Котовым и Э. П. Лебедевой (1977), Л. А. Животовским (1984), А. П. Царевым (1985, 2000, 2001), Л. Ф. Семериковым (1986) и рядом др., в том числе J. Hartwig (1892), G. Hegi (1908), Е. Silva-Tarouca (1913), S. Sokolowski (1931), E. Rohmeder, H. Schonbach (1959).
Внутривидовая изменчивость основных лесообразователей страны изучалась на основе накопившегося опыта методических подходов (Hartwig J., 1892; Schwerin F., 1906; Hegi G., 1908; Silva-Tarouca E., 1913; Филипченко Ю. A., 1927; Козубов Г. M., 1962a; Правдин Л. Ф., 1964; Rohmeder E., Schonbach H., 1959; Staszkiewicz J., 1961, 1963; Парамонов A. A., 1967; Завадский К. M., 1967; Альбенский A. В., 1941; Мамаев С. A., 1972; Ирошников А. И. и др., 1973; Хохрин А. В., 1977; Черепнин В. Л., 1970; Некрасова Т. П., 1959; Котов М. М. и Лебедева Э. П., 1977; Пчелин В. И., 1959; Коновалов Н. А и Пугач Е. А, 1978; Матвеева Р. Н., 1988; Хасанов Н. X., 1992 и др.
Однако нами применительно к лесным древесным растениям был разработан и предложен новый подход, основанный на выделении и учете маркерных признаков как качественной, так и количественной природы их происхождения. На необходимости использования данного подхода акцентировали внимание Н. В. Тимофеев-Расовский и А. В. Яблоков (1973) и С. С. Шварц (1980).
Из комплекса структурных признаков (к примеру, у сосны обыкновенной) изучались как качественные, так и количественные признаки – такие, как: общая высота и высота до первого живого сучка, диаметр на высоте груди, протяженность грубой корки, ширина, плотность, архитектоника и протяженность кроны, строение и окраска коры, объем ствола, угол отхода сучьев и их толщина, левизна-правизна шишек, длина и цвет хвои, окраска генеративных органов и ряд др.; учитывались и резко уклоняющиеся от стереотипа особи (к примеру, гроздешишечные, с "ведьмиными метлами", с "воротничковым" строением коры, "кипарисовидные" кроны, с полным опаданием хвои предыдущих лет, с короткой хвоей и целый ряд других отклонений).
Изменчивость шишек определялась по форме, массе, размерам (длина, ширина), окраске, форме апофизов на концах семенных чешуй; семян – по массе, цвету. Для определения цвета была использована шкапа цветов А. С. Бондарцева (1954), но ввиду того, что спектр признаков (в особенности качественных) в природе из-за наличия промежуточных и переходных форм гораздо шире и они обычно не укладываются в те регламентирующие ограничения в имеющихся методических пособиях, нами были введены новые промежуточные градации и дополнения (и к упомянутой шкале цветов А. С. Бондарцева в частности). Для определения левизны-правизны шишек была использована методика А. В. Хохрина (1970, 1977). При изучении структурных особенностей строения и архитектоники кроны в основу были положены термины общей систематики, ботанической номенклатуры, морфологии и анатомии растений и ряда других ботанических дисциплин (Кирпичников М. Э. и Забинкова Н. Н., 1977). По очищаемости ствола от сучьев деревья подразделяли на три категории: с хорошей (единичные сухие сучья по стволу у основания живой кроны), с плохой (сухие сучья по стволу составляют до половины длины ствола до кроны) и средней (сухие сучья по стволу расположены до 1/3 длины ствола под живой кроной). При описании структуры популяций фиксировали одновременно как качественные, так и количественные структурные признаки и их частоту в совокупности особей. Кодирование признаков осуществляли самыми различными способами, начиная от условных знаков и кончая индексами в позиционной, полупозиционной и непозиционной записях.
Способы решения задач сравнительного анализа фенофондов применены, начиная с графического сравнения до подключения методов машинного анализа на ЭВМ. При инвентаризации насаждений использовали традиционные в полевых исследованиях методы: маршрутно-рекогносцировочный и метод пробных площадей. Не исключался и метод массовой селекции со снятием всех интересующих параметров модельных деревьев и сбором растительных образцов. Были разработаны математические модели, алгоритмические схемы и пакеты прикладных программ ОТЭКС для ЭВМ предложенных в исследовании методов.
Экспериментальный материал анализировали различными методами сравнительного многомерного анализа (СМА) с подключением ЭВМ: таксономический и кластерный анализы, в том числе Q-метод и групповой коэффициент связи (ГКС), изотропный метод СМА, координатный метод комплексной оценки, метод на основе показателей, позволяющих оценить внутрипопуляционное разнообразие и степень сходств и различий сравниваемых насаждений по полиморфным признакам (Животовский Л. А., 1979, 1980, 1982). Анализ хода роста стволов проведен по принятым в лесной таксации методам (Семенюта Ф. И., 1968).
Достоверность выводов и результатов исследований обеспечена использованием значительного по объему экспериментального материала, необходимого числа повторностей при закладке опытных объектов, воспроизводимостью полученных результатов и критериями достоверности в различных лесорастительных регионах ареала древесных пород. Весь опытный материал обрабатывался методами математической статистики и СМА с разработкой математических моделей. Для оценки взаимосвязи признаков применялся корреляционный анализ, критерий t – Стьюдента и критерий Фишера, коэффициент наследуемости h2 и ряд других биометрических показателей; при оценке законов распределения – критерий Хи-квадрат (?2) При сравнении фенофондов и фенотипических структур использованы также показатели внутрипопуляционного разнообразия как среднее число вариаций признака ?, доля редких фенов h, показатель сходства популяций r, критерий идентичности популяций I, секториальные графики (векторные диаграммы) и ряд других статистических характеристик.
Широкий спектр применяемых методов анализа особей и популяций позволил отчетливо их "расшифровать" по комплексу признаков на определенные группы-кластеры и выявить среди них хозяйственно ценные формы для дальнейшего их использования как доноров семенного и вегетативного материала при создании постоянной лесосеменной базы на генетико-селекционной основе в тех или иных климатических зонах ареалов основных лесообразующих пород страны.
ГЛАВА 2 ФЕНЫ ДРЕВЕСНЫХ РАСТЕНИЙ
2.1. "ПОЛИМОРФИЗМ" ФЕНОВ
У древесных пород в качестве основных изучаются три группы признаков-маркеров: структурные (морфологические), отражающие особенности строения формы и размеров органов и их частей; функциональные (физиологические), характеризующие особенности физиолого-биохимических процессов, и биохимические, выявляющие вариации в содержании органических соединений в тканях (Яблоков А. В. и Ларина Н. И., 1985).
Н. И. Вавилов (1920) провел изучение качественных признаков на растениях, в основу которого он положил сравнение легко выделяемых и наследственно обусловленных дискретных вариаций признаков, вполне вписывающихся в понятие "фен". Накопившиеся сведения по выделению и изучению фенов древесных растений на сегодняшний день уже весьма обширные, и спектр их представлен категориями разных уровней – структурного, функционального и биохимического.
Л. Ф. Правдин (1964), в частности, в изменчивости морфологических и анатомических признаков сосны выделяет размеры, анатомическое строение, расположение смоляных каналов, аномалии в анатомическом строении центрального цилиндра хвои, изменчивость размеров, строения и окраски шишек, изменчивость репродуктивных органов (морфология и окраска "соцветий", изменчивость пыльцы, морфология и физические свойства семян, изменчивость массы семян, их цвета и цвета крылышек, качество семян и размеры семядолей). По изменчивости некоторых экологических особенностей и физиологических свойств сосны определена продолжительность жизни хвои, сезонная изменчивость пигментов хвои, фотосинтез и транспирация сосны разного происхождения, жирность семян, рост и продуктивность сосны, изменчивость кариотипа. Различает он также формы сосны по архитектонике кроны и форме ствола, строению грубой корки, размерам и окраске хвои, качеству древесины, окраске репродуктивных органов, строению шишек и цвету семян и зрелых шишек.
Авторы "Методики изучения внутривидовой изменчивости древесных пород" (1973) выделяют три группы признаков: а) структурные (27 признаков, начиная с класса роста дерева и заканчивая числом и формой хромосом); б) функциональные (9 признаков, начиная с энергии прироста в течение сезона и до общего количества ассимилируемой массы); в) качественные (6 признаков, начиная с масличности семян и до количества и состава фенольных веществ).
С. А. Мамаев и А. К. Махнев (1982, 1983, 19.88) выделяют стабильные и лабильные признаки. Причем к стабильным они относят окраску семян у хвойных, форму чешуй и апофиз шишек, окраску генеративных органов (у хвойных), форму плодовых чешуй (у берез), белковый и изоферментный спектры; к лабильным признакам – размеры и форму листьев, размеры семян, шишек и сережек (у березы), окраску и строение корки у березы, количество смоляных каналов в хвое, опушенность листьев у березы, уровень термофильности и продолжительность периода покоя семян, термостойкость органов и содержание антоцианов. Помимо стабильных и лабильных эти авторы выделяют и признаки промежуточного типа. Они отмечают, что многие структурные видовые признаки (форма листа, строение коры и ряд др.) одновременно являются признаками таксонов низшего ранга и характеризуют структуру популяций. То есть метод морфофизиологических (генетических) маркеров (фенов) включает в себя использование как качественных, так и количественных признаков.
С. А. Петров и др. (1982, 1983) констатируют, что одной из главных проблем фенетики древесных растений является проблема выявления элементарных вариаций (составляющих) тех или иных признаков – к примеру, у сосны обыкновенной: окраска пыльников, способность меристемы к регенерации в стерильной культуре, изменчивость монотерпенов, рост деревьев, морфологические и цитологические особенности их потомства, архитектоника кроны (пирамидальная, плакучая и др.); форма апофиза шишек (плоский, выпуклый, крючковатый), цвет семян; геномные мутанты, где в качестве "фена плоидности" выступает целый комплекс признаков ("комплексный фен"). Анализ роста дерева позволил выявить динамическую и стохастическую составляющие этого сложного признака. Частота встречаемости вторичных перетяжек и их локализация на плечах ядрышкообразующих хромосом также могут служить в качестве манерного признака-фена. В качестве своеобразных фенов может быть использована изменчивость и ряда других признаков – в частности, физиолого-биохимических (качественный состав монотерпенов хвои у сосны). Качественная обусловленность способности ткани к каллусогенезу (при определенных условиях среды) позволяет рассматривать это свойство как элементарный признак-фен. Таким образом, изменчивость какого-либо количественного признака можно "свести" к нескольким дискретным классам путем использования относительной изменчивости признака в многомерном анализе.
Л. И. Милютин (1978 – 1990) изучает популяции лиственниц по 14 морфологическим, анатомическим, физиолого-биохимическим и кариологическим признакам, причем 6 признаков из морфологических могут быть отнесены к фенам (цвет молодых женских шишек, форма края семенных чешуй, тип плоскости семенной чешуи, форма шишек, цвет молодых побегов и их опушенность). Маркерами для отдельных деревьев лиственницы могут также служить биохимические фены – в частности, изоэнзимные спектры ферментов (пироксидаза, эстераза и др.). Подводя итог многообразию фенов у хвойных пород, он среди морфологических фенов выделяет фены генеративных и вегетативных органов; кариологические, биохимические, анатомические фены также требуют дальнейшей систематизации для широкого их использования в фенетических исследованиях.
М. Н. Егоров (1983, 1985, 1990, 1997, 2001) предложил перечень качественных и количественных признаков на примере сосны обыкновенной для широкого их использования в качестве маркеров при анализе фенотипической структуры популяций, причем спектр фенов, наряду с морфологическими показателями, может быть дополнен физиолого-биохимическими, анатомо-гистохимическими и др. В частности, у сосны обыкновенной диагностические признаки могут быть представлены следующим перечнем:
а) качественные альтернативные признаки – цвет хвои (темно-зеленый, зеленый, желтовато-зеленый, зеленовато-желтый, синевато-зеленый); цвет коры (бледно-красновато-бурый, табачно-бурый, золотисто-желтый, желтый, бледно-желтый, кровяно-красный); цвет зрелых шишек (темно-дымчатый, зелено-серый, коричневый, рыжеватый, темно-серый, медово-желтый, зеленовато-серый, зелено-малахитовый, зеленый, охристый); цвет семян (черный, коричневый, бледно-сероватый, пепельно-серый, белый); цвет женских соцветий (кровяно-красный, травяно-зеленый, каштановый, оливко-зеленый); цвет пыльников (желтый, оранжево-красный); цвет крыла семени (терракотовый, коричневый, бледно-терракотовый); строение коры (чешуйчатая, пластинчатая, чешуйчато-пластинчатая, пластинчато-чешуйчатая); форма апофиза шишек (плоская, бугорчатая, крючковатая); отпад хвои (второго, третьего годов жизни); периодичность семеношения (каждый год, через 1, 2, 3 и т.д. года, вообще неурожайное); архитектоника кроны (яйцевидная, конусовидная, флаговидная, веретеновидная, шаровидная и др.). Не исключается при этом подбор и ряда других признаков (к примеру, левизна-правизна шишек и побегов, угол отхода сучьев и др.);
б) количественные (размерные) признаки: высота, диаметр, объем ствола и общая биомасса дерева, высота поднятия фубой корки, толщина коры, число ветвей в мутовке, длина хвои (короткая, "нормальная" и длинная); степень охвоенности побегов (густая, средняя и редкая), число шишек в мутовке (в том числе и гроздешишечные особи); длина, ширина и масса шишек и семян и целый комплекс других признаков, к которым применимы биометрические методы анализа.
А. В. Хохрин (1983) утверждает, что зеркальные, или энантиоморфные, признаки являются хорошими диагностическими признаками-фенами. У древесных растений с винтовым листорасположением и моноподиальным ветвлением ствол дерева образуется в двух зеркальных формах – левой (L) и правой (D). Частота встречаемости L- и D-форм у основных лесообразовате- лей – сосны, кедра, ели, пихты и лиственницы варьирует около равночисленного, т.е. как 0,5:0,5.
М. В. Придня (1977, 1983, 1985) у бука восточного выделяет прилистники и шиповидные образования купул, брохио-дромность и краспедодромность листа, лопастность и зубчатость околоцветника мужских цветков; у нордманновой пихты – цельность и выямчатость листа, выход язычка кроющей чешуи за пределы семенных чешуй женских шишек и отсутствие выхода (язычок в пределах кроющих чешуй).
А. И. Ирошников (1983, 1985) генетическую структуру популяций характеризует комплексом дискретных моногенных (реже полигенных), стабильных в онтогенезе (или на отдельных его этапах) признаков различных органов и структур древесных растений Число фенов у отдельных признаков колеблется, как правило от 5 – 9 до 40. Особенно много фенов выделяется по цвету и рисунку окраски семенных чешуй у шишек лиственницы сибирской и ели сибирской. В зависимости от задачи исследования фены могут объединяться в определенные группы. Элементарные признаки эффективно выявлять в области оптимума вида, в гибридных популяциях и среди потомства от контролируемых скрещиваний контрастных фенотипов. Информативность исследований генотипического состава природных популяций хвойных пород повышается при использовании не только альтернативных моногенных качественных признаков, но и признаков-маркеров, имеющих более сложную природу.
Л. Ф. Семериков и Н. В. Глотов (1983) для характеристики изменчивости использовали мерные и счетные морфологические признаки листьев и генеративных органов у дуба – плодоноса, плюски и желудей, что позволило им, в первую очередь, выделить на Кавказе 6 групп популяций, в Поволжье и Предуралье – 5 групп популяций дуба черешчатого.
А. В. Яблоков и Н. И. Ларина (1985) отмечают, что у древесных растений исследуются все три группы признаков: морфологические, физиологические и биохимические. Опыт работы палеонтологов с хвойными показал, что только на пыльце хвойных деревьев им удалось выделить 33 признака со 104 дискретными вариациями, подавляющая часть которых, по-видимому, может рассматриваться как фены. В древесине же палеозойских голосеменных растений – кордаитовьгх и хвойных – описано 39 признаков со 175 дискретными вариациями.
Б. А. Кравцов и Р. Г. Хлебопрос (1985) на примере лиственниц Сибири показали полезность ряда непараметрических процедур при фенетическом анализе природных популяций, предложив методику ранжирования фенов по их информационной ценности.
С. А. Мамаев, А. К. Махнев и Л. Ф. Семериков (1985) при выделении генетических резерватов в первую очередь рекомендуют учитывать показатели продуктивности (диаметр и высота деревьев, качественный состав и состояние древостоя, прямизна и очищаемость ствола от сучьев, устойчивость к разного рода заболеваниям, повреждениям и т.д.). Объективным совокупным показателем в данном случае может быть класс бонитета насаждения. Достаточно изученными маркерами являются признаки, характеризующие сбег деревьев и структуру кроны (форма, протяженность, обилие и размеры сучьев и угол их отхождения от ствола), окраску и строение коры, плодов и т.д. Некоторые из отмеченных признаков коррелятивно связаны с признаками, которые не поддаются непосредственной оценке (к примеру, строение и окраска коры отражают особенности текстуры и физико-механические свойства древесины). В итоге эти авторы указывают, что непосредственно выделение результатов осуществляется с помощью генетических маркеров.
Ю. Е. Булыгин (1985) при типизации структуры древесных растений рекомендует учитывать в опыте различные показатели признаков или свойств исследуемых лесных пород (из количественных признаков – диаметр ствола и его высоту, величину текущего прироста по высоте и диаметру и ряд др.; из качественных признаков – строение и цвет коры, зрелых шищек и семян и др.).
К. Буторина и Л. С. Мурая (1985) при сравнительной характеристике генотипов хвойных по данным их кариотипичес- кого анализа устанавливают значительный полиморфизм по частоте встречаемости и локализации вторичных перетяжек, а также по абсолютной длине хромосом набора у сосны обыкновенной и трех видов сосны с Кубы: Pinus cubensis, Pinus caribae, Pinus occidentalis.
A. Духарев (1985) при изучении генотипической структуры 20 популяций сосны обыкновенной на всем протяжении ее ареала на территории бывшего СССР использовал в качестве маркеров изоферменты, контролируемые 42 локусами. При этом система размножения хвойных, как и других голосеменных, такова, что дает возможность использовать одновременно и диплоидную (например, хвоя) и гаплоидную (эндосперм) фазы развития.
Ю. Н. Исаков (1985), рассуждая о природе исключительно высокого фенотипического полиморфизма березы карельской, отмечает полиморфность многих ее признаков, в том числе узорчатость древесины и толщину коры как представляющие хозяйственно ценные признаки. Ю. Н. Исаков и В. В. Иевлев (1985) при изучении изменчивости линейных приростов отдельных деревьев в выборках сосны обыкновенной выявили две составляющие роста дерева – детерминированную и случайную. Они заключают, что неслучайность проявления элементарных составляющих роста предполагает генетическую дискретность развития деревьев, т.е. наличие фенов онтогенеза. Они же, рассматривая генетическую обусловленность некоторых фенов хвои сосны, в гибридных и контрольных (от свободного опыления) семьях вьщеляют несколько групп фенов-маркеров по окраске хвои (с зеленой, желтовато-зеленой, желтой и зеленовато-желтой); по расположению хвои на боковых побегах (радиальное и гребенчатое). По их мнению, выделенные фены могут служить надежными маркерами генотипов адаптивных признаков.
А. А. Клявин (1985) при фенетическом анализе дуба черешчатого в Воронежском заповеднике исследует также количественные и меристические признаки: длина листа, ширина листа по лопастям, ширина листа по бухтам, число первичных лопастей, число вторичных лопастей, длина черешка, число дополнительных жилок, отношение ширины листа по лопастям к его длине, а также ширины листа по бухтам к его ширине по лопастям. Кроме того, для признаков 2, 3, 4, 5 и 7 значения параметров брались справа и слева от центральной жилки, а затем вычислялось отношение разности этих значений к их сумме.
А. И. Обыденников (1985), устанавливая генотипический состав лесных популяций, приходит к выводу, что расщепление признака числа семядолей в потомстве исходного материала у хвойных растений (на примере лжетсуги Мензиса), коррелирующего с изменением показателя интенсивности роста, соответствует биноминальному распределению по однолокусовой модели формулы Харди–Вайнберга.
С. А. Петров (1985, 1990) также отмечает, что распределение частот различных фенов в популяциях нередко является сбалансированным и подчиняется теоретическому распределению Харди–Вайнберга. Данные по сравнительному изучению метамерной и внутриполяционной изменчивости таких дискретных признаков, как окраска микростробилов, зрелых шишек, семян, побегов, хвои, опушение побегов, строение края семенной чешуи, обладают очень высокой метамерной стабильностью и, как правило, не связаны между собой и свободно комбинируются в самых разнообразных сочетаниях. Подобными характеристиками обладают и некоторые количественные признаки (точнее – их индексные оценки) – такие, как форма шишек, плодов, семян, листьев, метамерная изменчивость которых практически не проявляется, а значения коэффициентов повторяемости достигают очень высоких уровней, т.е. формирование таких признаков и их изменчивость в популяциях находятся под жестким генетическим контролем и поэтому также могут быть использованы в качестве признаков-маркеров генотипического состава популяций.
И. М. Пожидаева и др. (1985) с помощью цитологических критериев оценивали генгетическую структуру популяции сосны обыкновенной и анализировали характер прохождения мейоза при микроспорогенезе. При этом выделены типы нарушения хромосом (забегания, отставания, образование мостов, фрагментов, микроядер и др.), которые, естественно, накладывают свой отпечаток на формирование гамет. Статистический анализ данных показал неоднородность характеристик и дискретный характер распределения деревьев сосны по частоте нарушений, что указывает на возможность использования данного показателя в качестве маркера генетической конструкции особи, а их частотное распределение характеризует структуру популяции по данному признаку.
Ю. К. Подгорный (1985, 1990) предпринимает попытку рассматривать садовые формы древесных растений с точки зрения фенетики (описано свыше 1000 "садовых форм" у 300 видов древесных пород), представленные в основном морфологическими мутантами с резко измененной кроной (33 %) и размерами, формой и окраской листьев (51 %), измененным ветвлением и строением побегов (1 %), с измененной формой роста (9 %), с окраской и строением цветка или его частей (1 %), имеющих резкие отклонения от стереотипа. Проводя анализ структуры высотных популяций сосны крымской по фенам окраски микростробилярных колосков, он отмечает, что по окраске мужских колосков она разделена на две группы – красно- и зелено-колосковые, и данные признаки рассматриваются как нейтральные, не подвергающиеся непосредственному действию естественного отбора.
Е. А. Пугач (1985) предполагает изучать идентификацию признаков генеративных органов сосны обыкновенной по цвету семенной кожуры и крылышка, цвету и форме апофиза шишек и цвету мужских и женских стробилов, утверждая при этом, что совместное однообразное сочетание всех шести признаков возможно лишь у генетически родственных деревьев.
М. Г. Романовский (1985) при изучении групповой измен- чи^ти древесных растений по количественным признакам ре- ко^^ так называемые "признаки-соотношения" как разряд высот Гсоотношение высоты и диаметра ствола), допускающих обобщение разновозрастных и разнобонитетных выборок. Данный подход позволяет однозначно оценить структуру выборки без определения групповой принадлежности каждой отдельно взятой особи по ряду количественных признаков с непрерывным спектром изменчивости, что расширяет возможности использования фенетических методов.
В. А. Углов (1985) при анализе ландшафтно-экологической изменчивости частоты фенов формы семенных чешуй в популяциях ели в Рязанской области вьщеляют три фена формы семенных чешуй: Picea onbiculata (о), Picea unidentata (u), Picea bidentata (b). Существование явления ландшафтно-экологической изменчивости частот фенов, по мнению автора, имеет и прикладное значение в картографировании фенотипических структур природных популяций.
Р. Г. Шеверножук (1985) допускает возможность выделения фенов для ряда адаптивных признаков функциональной природы у древесных растений. Биоэлектрические реакции, наблюдаемые в популяциях лесных пород, предлагается разбить на определенные классы растений с различной способностью реагирования на внешние воздействия, которые, по мнению автора, можно считать фенами адаптивных признаков функциональной природы, характеризующимися следующими особенностями: они проявляются в особых условиях при инструментальном определении, склонными к флуктуациям в разные годы. Автором предлагается к применению термин "функциональный фен".
А. М. Шутяев (1985) при фенетическом анализе географических популяций дуба черешчатого по опушению листьев признает его одним из диагностических признаков и утверждает, что фен опушенности – это эволюционно лабильный признак. На основе изучения опушенности листьев у 60 потомств географических популяций ранней разновидности дуба, выращенных в ЦЧО, описаны география фена опушенности и структура популяций по данному признаку.
Г. Г. Гончаренко, В. Е. Падутов и В. В. Потенко (1989) при исследовании хвойных видов методом электрофоретического анализа изоферментов подчеркивают, что особенно перепективно применение методов генетики изоферментов в лесной селекции, так как длительная смена поколений у древесных не позволяет использовать традиционные методы генетики и селекции для улучшения хозяйственно-ценных признаков деревьев. Белки и ферменты, подобно зеленому и желтому цвету семян у Г. Менделя, находятся под контролем отдельных генов, однако, в отличие от признаков в опытах Г. Менделя, ферменты практически всегда экспрессируются кодоминантно. Это означает, что мы можем выявить фенотипические (видимые) различия между растениями, которые имеют генотипы АА, Аа, аа при использовании метода электрофореза. Изоферменты (наследуемые фракции ферментов) обладают целым рядом преимуществ по сравнению с другими классами наследуемых генетических маркеров. Несколько изменчивых альтернативных признаков найдены и у древесных хвойных видов (форма семенной чешуи, цвет семенных крылаток, размеры и форма семян; имеются и другие морфологические вариации с вредным или летальным эффектом – карликовость, альбинизм). Однако определить, контролируется ли тот или иной признак у древесных конкретными генами или зависит от случайных обстоятельств, является пока задачей очень сложной. Другой класс изменчивых признаков у хвойных был обнаружен по монотерпенам, причем оказалось, что высокий и низкий уровни монотерпенов контролируются генами, но, в отличие от морфологических маркеров, различия по монотерпенам найдены только между популяциями, а не между отдельными деревьями, и в связи с этим авторы находят, что вряд ли монотерпены можно рекомендовать в качестве удобных генетических маркеров в селекции хвойных.
А. И. Видякин (1990) предложил индексно-фенетический метод выделения популяций сосны обыкновенной по количественным признакам, и непрерывная индивидуальная изменчивость индексов за счет этого (путем разделения пределов изменчивости каждого индекса на классы) преобразуется в дискретную, и в этом смысле данные вариации отвечают понятию фена. Для сосны обыкновенной выделены цветосеменные фены-маркеры генотипического состава популяций, причем границы популяций, устанавливаемые на основе многих индексов и цветосеменных фенов, совпадают.
Г. Г. Гончаренко и др. (1990) при генетической и фенетической дифференциации в видовых формах ели европейской находят дискретные фенотипические вариации по строению шишек форме семенных чешуй, цвету и форме крылаток, одновременно указывая, что они не являются определяющими для выделения видовых форм ели европейской.
В. П. Демиденко и В. М. Урусов (1990) при изучении популяционной структуры сосны на Алтае ее изменчивость в данном регаоне определяли через географические закономерности распределения форм по строению апофизов шишек, величине и среднему количеству семенных чешуй, массе семян, продолжительности жизни хвои, цвету мужских стробилов перед цветением.
В. И. Долголиков (1990), рассматривая перспективы селекции широко- и узкокронных форм ели в природных популяциях Ленинградской области, отмечает, что сложившееся мнение о селекционной значимости диагностического признака – ширины кроны не однозначно, но он не должен игнорироваться в селекции древесных растений.
Ю. Н. Исаков и др. (1990), изучая полиморфизм древесных видов по уровню самофертильности (самофертильные, самостерильные, частично фертильные особи), заключают, что учет частотного распределения деревьев в выборках различных популяций по уровню самофертильности позволяет дифференцировать подход к выбору методов генетического анализа и планировать оптимальные схемы селекции.
Н. Е. Косиченко и др. (1990) отмечают появление признаков узорчатости древесины в потомстве березы при гамма-облучении семян, сопровождаемом аномальными изменениями структуры проводящих тканей и стебля, обусловливающими развитие высокодекоративной узорчатой древесины, которая может быть представлена также в ранге диагностического признака-маркера.
В. Е. Кулаков (1990), выделяя природные популяции кедра сибирского, заключает, что пока не установлены надежные признаки-маркеры, определяющие генетическое различие популяций, и наибольший интерес представляют признаки: масса 1000 шт. семян и урожай их с одного дерева. Оказалось, что различие их внутри одного массива – лишь 4,3 %, между массивами – от 13 до 27 %, хотя лесоводственно-таксационная характеристика их аналогичная. Это дает повод предположить, что последнее генетически обусловлено
Я. Ларионова (1990), анализируя аллозимную изменчивость популяций лиственницы, приводит данные электрофоретической изменчивости 6 ферментных систем в гаплоидной и дишлойдной тканях семян лиственниц Сукачева и сибирской из 10 популяций, которые показали, что многие из обнаруженных электрофоретических вариантов этих ферментных систем являются продуктами аллелей отдельных генных локусов и вполне могут быть представлены в качестве фенов для изучения изменчивости лиственниц, и использование биохимических фенов значительно расширяет возможности исследования изменчивости популяций хвойных пород.
М. Максимов и И.В. Кошовский (1990), исследуя фенотипическую структуру популяций сосны обыкновенной в ЦЧО по составу эфирных масел хвои, отмечают, что доминирующими компонентами являются ?-пинен и ?3-карен, синтез которых находится под генетическим контролем, и данные об их содержании можно использовать для изучения структуры популяций сосны в качестве фена.
А. К. Махнев (1990), используя признаки-маркеры при исследовании природных популяций древесных растений в экстремальных условиях, к их числу в первую очередь причисляет относительно стабильные и разного рода аномалии вегетативных и генеративных органов.
Е. Н. Муратова (1990), исследуя полиморфизм ядрышкообразуших хромосом у представителей семейства сосновых, предлагает использовать ряд кариологических признаков в ранге фенов, которые могут служить маркерами на уровне вида, популяции и в некоторых случаях – отдельной особи. У изученных видов сосны имеется от 4 до 7 хромосом со вторыми перетяжками, Лц-2–3, Е-5, П-4. Максимальное число ядрышек в интерфазных ядрах соответствует количеству хромосом со вторичными перетяжками.
Е. А Пугач и др. (1990) для оценки генотипического состава популяций сосны используют морфологические признаки генеративных органов, остающиеся неизменными в процессе онтогенеза, по их качественным характеристикам (микро- и макростробилы, цвет и строение зрелых шишек и их составляющих элементов: чешуек, крылышек и семян), и они, в свою очередь, служат надежными признаками-маркерами при изучении генотипического состава популяций.
Н. Ф. Храмова и В. П. Автономов (1990) считают, что по числу межмутовочных ветвей у взрослых особей ели сибирской можно дать предварительную оценку потенциального роста плюсовых деревьев в разновозрастных популяциях, где благодаря их фенотипическим особенностям оценка по прямому признаку иногда затруднительна. По числу боковых почек однолетнего побега можно также дать предварительную оценку роста отдельных полусибсовых семей ели сибирской.
И. Чернодубов (1990), наряду с В. М. Максимовым и И. В. Кошовским (1990), также утверждает, что в качестве признаков-фенов, генетически закрепленных, могут быть использованы ?3-карен и ?-пинен эфирных масел хвои сосны обыкновенной.
К. Ширнин (1990), рассматривая феногенетические аспекты исследования признаков древесины у лесных пород, отмечает, что с позиций фенетики годичные слои являются квантами морфогенеза, ежегодными метамерами, основными гомологичными структурными единицами древесного организма, характеризующими интегрально продуктивность и качество популяций и биотипов; сюда же следует отнести цвет и тип трещиноватости коры, форму кроны, время цветения и листораспускания и др.
М. М. Котов (1997) пишет, что формы древесных растений можно сгруппировать по признакам. В частности, биологические формы – это растения одного вида, различающиеся по интенсивности роста, конкурентной способности, возрасту возмужалости, особенностям половой дифференциации, опыления и семяношения, долголетию; к морфологическим формам по аналогии можно отнести узко- и ширококронные растения, редко- и густоохвоенные особи, красно- и желтопыльниковые, черно- и коричневосеменные, с желто-оранжевой и красновато-коричневой окраской тонкой коры, с плоским и крючковатым апофизом шишек, серо- и коричневошишечные, толсто- и тонковетвистые, с плотной и рыхлой древесиной. Физиологическими формами следует называть засухоустойчивые и чувствительные к засухе, требовательные к плодородию почв и мирящиеся с их бедностью, светолюбивые и сравнительно теневыносливые, с высокой и низкой продуктивностью фотосинтеза растения. Фенотипическое проявление таких признаков, как пол растений, окраска цветков и стробилов, окраска семян и крылышек, форма апофиза шишек и форма семени, плода и шишек и ряда других, не зависит от условий среды, а связано с определенным этапом онтогенеза. Внутрипопуляционная изменчивость по таким признакам также оценивается фиксацией растений с тем или иным фенотипом, вычислением участия растений с разными фенотипами в популяции, составлением гипотезы о характере наследования признака и подтверждением или опровержением гипотезы методом Хи-квадрат.
Из приведенного перечня признаков видно, что подавляющее большинство их может быть использовано в качестве маркерных признаков-фенов в структурном анализе популяций древесных растений. Вопрос лишь заключается в том, как правильно их "вычислить" в ранге маркерных признаков-фенов и целесообразно и эффективно с генетико-селекционной точки зрения использовать в научном анализе.
2.2. КАТАЛОГИЗАЦИЯ ФЕНЕТИЧЕСКИХ ПРИЗНАКОВ
Одной из основ исследований в области фенетики популяций является выбор морфологических характеристик, подлежащих учету в природных группировках особей, – фенов и фенокомплексов (а также таких признаков и их вариаций, которые соответствуют этим показателям). В данный момент одной из причин, сдерживающих широкое распространение фенетических исследований, оказываются трудности методического характера, связанные с выделением и описанием таких характеристик (Яблоков А. В., 1985).