Все указанные явления можно отнести к явлениям приспособления организма к окружающей его среде. Сюда же относятся и те случаи, когда внезапное выставление на простор подроста, находившегося под пологом насаждения, заставляет его тоже болеть или даже преждевременно отмирать. Народ хороша отметил явление засыхания елового подроста при его внезапном выставлении на простор таким образным выражением: подрост, мол, пугается. На самом деле, хвоя ели этого подроста, родившаяся в условиях большой тени, построена по теневыносливому типу и потому неспособна работать на сильном свету; кроме того, обладая более тонкой кутикулой и эпидермисом, она не в состоянии защитить свои органы от излишнего испарения, и действительно, хвоя на просторе очень часто у такого подроста желтеет и опадает. В ближайшие годы такое дерево, если не вся хвоя у него отпала, постепенно заменяет старую хвою новой, которая, залагаясь на свету, обнаруживает иное анатомическое строение и является приспособленной к новой окружающей среде. Когда такой подрост переболеет, т. е. приспособится к новым условиям жизни, тогда он начинает усиленно трогаться в рост, , и мы говорим о нем как о подросте благонадежном, как о подросте оправившемся. Степень прежнего угнетения подроста является решающим моментом того, оправится он или нет, точно так же, как и степень внезапности выставления его на простор. Само собой разумеется, что решающую роль в этом играет также и порода, степень ее пластичности, и климат, и почва; чем благоприятнее последние для данной породы, тем подрост выносливее, тем он легче оправляется, и наоборот. Новые условия простора вызывают, как нам отчасти стало известным, и резкие перемены то в составе живого покрова, то даже в составе будущего лесного поколения; вместо тенелюбов, растущих под пологом леса, поселяются светолюбивые травы и такие древесные породы, которые не боятся ни инсоляции, ни излучения, ни конкуренции буйной травяной растительности, как-то: береза или осина. Происходит иногда так называемая смена покрова и смена пород, как результат приспособления других организмов к новым изменившимся условиям жизни.
На особенностях внутренней среды леса и на ее экологическом значении я с удовольствием останавливаюсь несколько дольше в виду того важного значения, какое придаю как этой главе, так и всему отделу, имеющему в виду разъяснить биосоциальную природу лесных сообществ. Как этот отдел, так и следующий за ним, имеющий в виду научное освещение факторов лесообразования, как я уже упоминал, имеет для нас первостепенное значение, как фундамент для всей остальной постройки лесоведения. Дальше мы будем изучать законы роста леса, его возобновление и распространение, а также метаморфоз лесных сообществ, или так называемую смену пород, и, наконец, систематику, или так называемую типологию леса, где, однако, изучение различных форм или типов леса будет тесно связано с исследованием их свойств, с изучением их отправлений. И вот здесь этот громадный отдел – от роста леса до систематики включительно – не может быть понят без основательного изучения биосоциальной природы леса и факторов лесообразования. После этого краткого отступления позвольте вернуться к нашей основной теме.
Мы уже упоминали, что внутренняя среда леса устраняет светолюбивую растительность, защищает самосев от излучения и лишней инсоляции; теперь в дополнение нам нужно найти ответ к такому вопросу: может ли подрост под пологом испытывать недостаток влаги и даже погибнуть от этого? Прежде чем ответить на этот вопрос, обратимся к другому случаю, когда самосев появился возле леса где-нибудь на открытом месте; предположим затем, что он не страдает от заморозков, и обратим наше внимание на его отношение к влаге. Влаги в почве и грунте открытого места, как мы уже видели, будет больше, чем под пологом леса, и с этой точки зрения подрост на подобном месте без сомнения лучше обеспечен влагой; но, во-первых, для него тут может явиться конкурентом травянистый покров, буйно развивающийся в подобных местах; во-вторых, здесь сильно развиваются испаряющие факторы, т. е. тут дает себя знать ветер, наличествует большая сухость воздуха, и поэтому сама почва, помимо покрывающей ее растительности, здесь сильнее испаряет влагу. И вот, несмотря на большее богатство водой подобных открытых мест, подрост в подобных случаях очень часто, особенно в сухих областях и на сухих почвах, как показали наблюдения и опыты, отмирает вследствие засухи как в одиночном стоянии, так и в группах, притом без какого-либо следа повреждения его растительными или животными паразитами и без каких-либо помех в почве, вроде твердых прослоек ортштейна. В данном случае происходит усыхание подроста от нарушения баланса влаги – оттого, что расход превышает приход, оттого, что растение, несмотря на наличность влаги в почве, не в состоянии удовлетворить громадные запросы на влагу в силу развития на таких местах испаряющих факторов. Ранней весною на открытых местах могут засохнуть всходы еще от другой, но аналогичной причины, именно: могут быть дни с сильной инсоляцией на еще замерзшей почве, и тогда опять-таки влага не может поступить в растение в достаточном количестве, вследствие чего самосев гибнет. Посмотрим теперь, как обстоят дела с отношением подроста к влаге под пологом леса. Здесь прежде всего редко бывает замерзшая почва, а если таковая и бывает зимою, то к весне под влиянием снежного покрова она оттаивает, но если бы даже и этого не произошло, то замерзшие слои здесь не представляют той опасности, какой они являются на открытом месте, так как самосев под пологом сильно защищен от инсоляции и, кроме того, все испаряющие факторы под пологом леса сильно ослаблены. Теперь посмотрим, как дело обстоит в течение вегетационного периода. Многие полагают, что подрост под пологом леса гарантирован от засухи, но это не так, – и здесь, как и на открытом месте, он может отмирать от недостатка влаги, но только по другой причине. Здесь, правда, нет сильно испаряющих факторов, но зато в самой почве и грунте леса меньше влаги, и по мере развития дождливого периода, и по мере того, как мы будем передвигаться к концу вегетационного периода, влаги под влиянием конкуренции материнских корней будет становиться все меньше и меньше, так что возможно, и на самом деле это бывает, что действительно в почве нарушится баланс влаги, но уже не под влиянием усиленного расхода, а в силу недостаточного количества воды в почве и грунте. Такие явления, как мы увидим потом, часто наблюдались в сухих сосновых борах в засушливом климате нашей лесостепи.
Значение внутренней среды легко показать также на тех явлениях, которые происходят при внесении в состав лесного сообщества или другой породы или подлеска. Остановимся на последнем случае и предположим, что мы внесли искусственно или что само собой под пологом другого леса возник еловый подлесок. Мы будем иметь два случая: дубовый лес с обычным для него подлеском из лиственных пород и кустарников и такой же дубовый лес с еловым подлеском. Какая же разница будет в экологии или, в частности, в свойствах внутренней среды этих двух, теперь уже разных, лесных сообществ? Поселение ели прежде всего уменьшит приток осадков; вместо 20%, которые, вероятно, дубовый лес задерживал на своих кронах, молодой и густой ельник дополнительно задержит еще по крайней мере 40%, и уже от одного этого почва прежнего дубняка станет суше, и верхний ярус дуба уже от этой одной причины станет испытывать недостаток влаги. Затем поселившийся ельник понизит и приток тепла, или среднюю температуру почвы, почва станет холоднее, что для дуба, как для породы теплолюбивой, совершенно некстати. В-третьих, ель начнет складывать свой мертвый покров и в связи с климатическими условиями, ею создаваемыми, она начнет мягкий дотоле перегной превращать в кислый гумус, в котором жолуди плохо прорастают. В-четвертых, этот кислый перегной начнет усиленно выщелачивать почву, т. е., иными словами, сильно обеднять ее основаниями, и прежде всего перегнойные кислоты удалят известь, стоящую на страже выщелачивания почвы; обеднение же почвы опять-таки некстати для дуба – породы, требовательной к зольным элементам почвы. В-пятых, еловый подлесок, в сильной степени поглощая солнечные лучи, в сильной мере погасит тот свет, который светолюбивый полог дуба пропустил бы сквозь себя, и в силу этого светолюбивые дубовые всходы не в состоянии будут прозябать в полумраке под пологом ели. Но еще раньше, чем начнет плодоносить верхний полог дуба, как показали лесоводственные опыты, наблюдения, исследования и сама практика, сам дубовый полог начнет страдать от перечисленных влияний елового подлеска, именно в нем начнут обнаруживаться такие болезненные явления, как усыхание вершин и падение прироста. Недаром сложилась такая поговорка, что ель для дуба – волк, а бук, наоборот, – нянька дуба или мамка его; дело в том, что буковый подлесок под дубовым лесом не обнаруживает тех зловредных влияний, какие свойственны ели, в особенности на почвах сухих и бедных. Ель и дуб – породы двух разных ботанико-географических областей, и при смешении их происходит между ними та естественная борьба, которую мы можем наблюдать в пограничных районах Querceiuma и Piceetuma.
Мы упоминали уже о законе естественного изреживания насаждений; по мере хода изреживания от молодняка (чащи) до возраста спелости изменяется и внутренняя среда насаждения; изреживаясь, насаждения становятся более доступными осадкам и лучистой энергии; по мере же увеличения тепла и света кислый перегной приобретает более мягкий характер, улучшаются физические свойства почвы благодаря большему доступу осадков, с одной стороны, и уменьшению числа дерев, с другой; по мере изреживания насаждения увеличивается и влажность почвы и грунта. И вот как раз ко времени наступления возмужалости насаждения, когда оно начинает приносить обильно семена и условия для их прорастания, в особенности для дальнейшего развития всходов, становятся лучше, т. е. самосев спелых насаждений, благодаря изменившейся внутренней среде, хотя и будет испытывать конкуренцию со стороны материнских организмов, но в меньшей степени, чем если он появился в возрасте жердняка или в возрасте приспевающем. Тем не менее и в возрасте спелости конкуренция будет еще так велика, что на самосеве скоро скажется гнет материнского насаждения, и мы будем получать формы угнетенного подроста. В окнах, в небольших прогалинах, вдоль опушек условия внутренней среды будут более благоприятны, и потому там количество и качество самосева и подроста будут иные.
Не касаясь роли леса как климатического фактора в отношении влияния леса на ближние и дальние окрестности, мы еще раз на минутку остановимся на нем как одном из факторов климата, ограничивая, однако, его роль только тем пространством, которое находится под его пологом. Из всего вышеизложенного мы можем свести указанную его роль к следующим пунктам: лес под своим пологом делает климат суше в отношении количества осадков, но вместе с тем под своими кронами делает воздух влажнее и холоднее, здесь же он создает более спокойную атмосферу и сокращает вегетационный период; вместе с тем лес уменьшает под своею сенью доступ света и уменьшает амплитуды температуры. Все сказанное не дает возможности назвать климат под пологом леса каким-либо определенным термином, имеющимся в климатологии; уменьшение количества осадков делает климат суше, переносит как бы данное пространство в более сухой климат, но большая влажность и меньшие амплитуды температуры переносят данное место в более влажный и морской климат; таким образом, в конце концов климат под пологом леса имеет свой самобытный характер, или, точнее, таковым обладают те отклонения от климата данной местности, которые вызываются способностью леса претворять занятую им среду.
Теперь в таких же кратких словах нам остается охарактеризовать роль леса как почвообразователя. В этом отношении надо прежде всего указать на те три существующих момента, которые определяют его роль как почвенного фактора. Это, во-первых, сама совокупность деревьев с ее пологом наверху, с корневой системой внизу и со своей физиологической деятельностью; во-вторых, подстилка и процессы ее разложения и, в-третьих, те метеорологические условия, какие создаются под сенью леса. Лес прежде всего изменяет режим почвенной влаги; как он это делает, мы уже видели и потому останавливаться на этом не будем; во-вторых, он изменяет тепловой режим занятой почвы,- – она становится, как мы видели, в среднем холоднее, и амплитуды температуры ее уменьшаются; в-третьих, лес, обедняя своей корневой системой слои грунта в отношении зольных веществ, обогащает ими самые верхние горизонты почвы; в-четвертых, он благодаря кислому выветриванию более или менее сильно способствует выщелачиванию почвы, давая место проявлению так называемого подзолообразовательного процесса; в-пятых, по мере развития этого процесса происходят не только химические изменения, но и меняются физические свойства, а также структура почвы; в-шестых, наконец, всякий лес способствует сохранению земной поверхности от процессов развевания, сноса и смыва.
Можем ли мы лес рассматривать как средство наполнения и сбережения влаги? Для ответа на этот вопрос необходимо прежде всего принять во внимание такое расчленение: надо различать лесные массивы, с одной стороны, куртины, полосы и ленты леса – с другой; гидрологическая роль первых резко отличается от таковой же вторых. Лесные массивы, во всяком случае, не являются ни накопителями, ни сберегателями почвенной и грунтовой влаги, и если леса имеют определенное положительное влияние или на источники, или на режим рек, то происходит это от другой причины – от способности леса держать как бы под прикрытием земную поверхность. [1 См. сноску на стр. 224] Другое дело – отдельные куртины и полосы леса, которые, собирая благодаря понизовкам в своих недрах большие снежные скопления и благодаря относительно малому расходу влаги, могут при определенных условиях как рельефа, так и их собственной конструкции иметь положительное гидрологическое влияние на окрестности, защищая их от ветра, препятствуя сносу снега, способствуя более равномерному его залеганию и т. п.
В виде заключительного аккорда к этой главе я снова напомню о сложности и связанности всех явлений жизни. Мы видели, что по мере изреживания насаждения с большим доступом света неразрывно связан и больший приток тепла, а вместе с тем и больший доступ осадков; эти факторы отражаются на изменении характера или типа подстилки, а это обстоятельство в свою очередь имеет определенные полезные воздействия на почву; с другой стороны, все это происходит благодаря уменьшению числа стволов, следовательно, уменьшению конкуренции, что в свою очередь увеличивает влажность почвы, и т. д. и т. д. Вот перед нами первый ряд взаимозависимостей, первый ряд связей; но затем мы видейи, что эти коллективные влияния полога имеют и свои плюсы и свои минусы для жизни, ну хотя бы подроста, т. е. под пологом мы наблюдаем и явления защитного и покровительственного характера и явления иного порядка, характера конкуренции или соперничества, и опять-таки эти два рода явлений друг от друга неотделимы, они связаны друг с другом, хотя, правда, можно регулировать эти соотношения, можно путем прореживания леса ослабить факторы конкуренции, но изъять их из обращения все-таки нельзя, они всегда будут налицо вместе с факторами защитного характера и всегда будут неотделимы друг от друга, как две стороны медали.
Вот второй ряд связанностей который не стоит как-либо особняком, а всегда находится во взаимоотношении с первым рядом; здесь мы собственно констатировали отражение на растениях, живущих сообществами, связанных явлений первого ряда. Теперь, если обратиться к третьему ряду, именно к факторам лесообразования или, в частности, к факторам образования внутренней среды, то это будут, как мы знаем уже, внутренние свойства пород, географическая среда и сами социальные условия, т. е. будет ли насаждение простое или сложное, густое или редкое, чистое или смешанное и т. д. Эти три рода факторов, как мы увидим в следующем отделе, опять-таки взаимно связаны друг с другом, и вместе с тем они образуют ту или иную внутреннюю среду. Таким образом, мы видим как бы три ряда явлений, в каждом из которых отдельные явления, как звенья одной цепи, связаны друг с другом, вместе с тем все эти три ряда, или три цепи, связаны в свою очередь друг с другом. Таким образом, мне хотелось лишний раз показать всю сложную ткань жизни, всю сложную ткань внутреннего жизненного механизма леса и тем противодействовать всяким односторонним направлениям, которые, избрав тот или иной факт, стараются все явления привести к нему и все объяснить исходя из него. Живая сложная жизнь обычно, однако, не повинуется ни доктринерству, ни догматизму, и тот, кто хочет не только понять ее, но и овладеть ею, влиять на нее, регулировать ее проявления, тот не должен терять чувства действительности, не должен терять из виду сложность, многогранность, взаимную обусловленность и связанность жизненных явлений.
Мы в дальнейшем изложения, когда остановимся на свойствах чистых и смешанных, простых и сложных, одновозрастных и разновозрастных лесных сообществ, когда приступим к ознакомлению еще с некоторыми особенностями роста леса, его возобновления, его динамики и смены пород, а также с типами насаждений, будем иметь бездну случаев иллюстрировать важность вышесказанного в ряде конкретных примеров.
О РОСТЕ НАСАЖДЕНИИ В СВЯЗИ С ЯВЛЕНИЕМ БОРЬБЫ ЗА СУЩЕСТВОВАНИЕ МЕЖДУ ДЕРЕВЬЯМИ
Имея в виду остановиться в дальше предстоящих главах на таких природных свойствах леса, как его пластичность, его способность лепиться в разнообразные формы в зависимости, от внешних условий, в связи с внутренними биологическими свойствами составляющих его элементов и под влиянием человека, обратимся предварительно к некоторым деталям, относящимся к другому свойству леса, образующему другую коренную черту его биологической физиономии, – к борьбе за существование, которая, как закон жизни, царит в лесу. Вполне уместно поставить такой вопрос: в чем же она выражается или в чем она должна выражаться, для того чтобы это образное выражение не было звуком пустым, а отвечало бы действительному ходу вещей? Мне кажется, что на этот вопрос дает превосходный ответ наш известный лесовод Я- С. Медведев. Если какой-нибудь орган дерева или все деревцо погибает под влиянием заморозка, молнии, ветра и тому подобных неблагоприятных влияний среды, то такие явления представляют собою проявление борьбы растений с внешней природой; но тогда, когда такая же потеря частей или всего деревца есть результат отенения (т. е. недостатка света) соседним более мощно развитым организмом или есть результат недостатка влаги, отнятой более сильным соседом, то это – борьба за существование дерев друг с другом. На какой-нибудь единице площади помещается определенное число растений известного возраста; с увеличением последнего будут увеличиваться я размеры отдельных деревец, а стало быть и их потребности; вначале изолированные, организмы, постепенно разрастаясь, придут в соприкосновение друг с другом как в кронах, так и в корневых системах. С этого момента мы будем наблюдать следующие явления: размеры и потребности деревец будут увеличиваться, а площадь останется без изменения; вширь раздаваться им можно только до известного предела, а потому при завоевании пространства им остается расти вниз и вверх; вниз до меньшего предела, вверх – до большего. В таком положении приобретает большое значение быстрота проста стволов, а также, по всей вероятности, и корневой системы; когда насаждение сомкнётся, т. е. в момент, когда оно образует сообщество оно однородно, так как различная индивидуальная сила роста у одного и того же вида, у одного и того же экземпляра не успела еще проявить себя в социальном отношении.
Если бы деревья были изолированными растениями, то индивидуальная сила роста, полученная по наследству от материнского организма и различная у разных индивидуумов, никаких особенно заметных последствий за собою не повлекла бы; но при совместном росте, когда в борьбе за пространство можно его; попользовать только вверх и вниз, а затем уже, после победы над соседом, и в сторону, различная индивидуальная сила «роста приобретает коренное значение, являясь первым и самым сильным (моментом в расслоении первоначально однородного сообщества и могучим фактором в борьбе за существование деревец друг с другом.
Те деревца, которые наделены большей индивидуальной силой роста, перерастают своих соседей и тем самым становятся в лучшие условия для использования тепла и света, и тем самым в то же время создают неблагоприятные условия для перегнанных ими в росте и отененных ими соседей. Отставшие экземпляры принуждены довольствоваться меньшим количеством света, что в свою очередь будет плохо отражаться на развитии других питающих органов корневой системы, а последняя, все хуже и хуже развиваясь, не может, конечно, содействовать энергичному развитию кроны, и таким образом устанавливается круг, который будет все более и более ухудшать питание и обессиливать отставшее деревцо. Эта борьба, как это давно отмечено и доказано в лесоводстве, не проходит бесследно и для победителей; последние на свободе, вне конкуренции других, несомненно развились бы лучше, как это доказывают те опыты в лесоводстве с прореживаниями, когда ограничиваются удалением одних только угнетенных классов, т. е. побежденных в борьбе за существование, и тем не менее получают более или менее сильный прирост в господствующих классах. По мере хода борьбы положение господствующих будет улучшаться, положение отставших, наоборот, ухудшаться; последние, все хуже и хуже развиваясь, попадут в разряд угнетенных, или заглушённых, т. е. очутятся под общим пологом, где прозябанию их поставлены различные границы, в зависимости от климатических особенностей местности и степени теневыносливости породы. Все весьма многочисленные исследования по этому предмету в лесоводстве, все без исключения так называемые опытные таблицы, отрывки из которых были приведены в начале книги, в главе II, неоспоримо доказывают факт наличности борьбы за существование и факт его постоянства до самого высокого возраста.
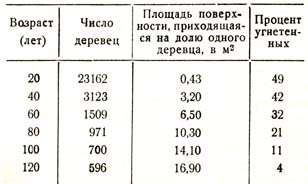
На основании данных, имеющихся в опытных таблицах, можно показать, как велик бывает объем пространства или площадь земной поверхности, занятой одним деревцом в различные возрасты насаждения. Для ели, например, мы имели следующие данные (табл. 43):
Таблица 43
На роль индивидуальной силы роста было уже отчасти указано выше; по всей вероятности, по мере большего изучения этого процесса в лесу, на который Дарвин почти не обратил внимания, но который с точки зрения дарвинизма обещает принести обильные плоды, будет больше разъяснена и роль индивидуальности растений в борьбе их друг с другом не только в отношении быстроты роста, но и других жизненных свойств, определяющих социальную жизнь растений. Точно так же и борьба деревец друг с другом ведется не только из-за света, а и из-за почвенной пищи; но последние моменты меньше скрыты и разъяснены. Вне всякого сомнения также, что в ходе борьбы за существование играет роль не одна только индивидуальная сила роста, но и случайные причины, благодаря которым могут погибнуть господствующие экземпляры и дать место, таким образом, угнетенным, которые, как показывает опыт, могут постепенно оправиться. Нет сомнения также, что значение в этом ходе борьбы за существование может иметь неполная однородность почвенной среды, хотя бы она и казалась однородной во всей своей толще, но так как индивидуальная сила роста может быть наблюдаема и в условиях совершенно одинаковых, как это проверено при опытах в искусственных средах, то она является началом более общим и первичным, а потому и первостепенным по сравнению с второстепенными, наслаивающимися, несколько лишь видоизменяющими основную картину влияниями неоднородностей среды и разного рода случайностей.
То обстоятельство, что угнетенные деревья при известных условиях, при устранении господствующих, могут оправляться, не может быть аргументом против индивидуальной силы-роста. Угнетенные деревья – не абсолютно худшие деревья, а лишь относительно менее приспособленные к жизненной обстановке данного места при наличности борьбы за существование; наличность угнетенных классов не доказывает, что эти деревца или эти породы, в случае состязания нескольких видов между собою, нежизнеспособны вообще; нет, они могут жить в изолированном состоянии, в парках, садах, при устранении борьбы за существование с другими, более сильными особями, наделенными в большей мере теми ценными качествами, которые необходимы в борьбе древесных растений друг с другом и которые, кстати замечу, и выработались не только под влиянием местного климата и почвы, но под непосредственным влиянием борьбы за существование; орудия для борьбы, иначе говоря, точились и вытачивались в самой борьбе друг с другом; среда влияла, как увидим ниже, на борьбу, но и борьба углубляла и усиливала значение условий местопроизрастания. Угнетенные деревья – относительно нежизнеспособный элемент только в лесу, предоставленном самому себе; если же устранить искусственно, как это иногда и делается, господствующие элементы, как, например, при прореживании по способу Борггреве, то такие деревца могут оправиться, пойти в рост, возмужать и благополучно создать потомство; частью они представляют собою в лесу как бы резервный фонд на случай, когда какая-нибудь случайность уничтожит в том или другом месте господствующий экземпляр, – освободившееся пространство займет «.кандидат на угнетение» (III класс по Крафту), а за отсутствием его – и сильно заглушённый экземпляр, если он не вполне еще потерял способность оправляться, или если перемена не наступает слишком внезапно; но обычно угнетенные деревья обречены на вымирание и на бесплодие, как на это было уже обращено внимание в начале книги в главе «Плодоношение разных классов господства. Естественный отбор», о чем еще будет подробно говориться в следующей главе. Благодаря именно обстоятельству, что угнетенные классы обречены на бесплодие, – в возрасте возмужалости каждого насаждения плодоносят преимущественно господствующие экземпляры – победители в жизненной борьбе, а не побежденные, т. е. в отношении размножения заявляют о себе наиболее приспособленные ко (всей обстановке, создаваемой в каждом данном месте лесом, а не менее приспособленные. Борьба за существование, о (Которой идет речь, которая постоянно составляет коренную черту леса и которая великолепно доказана лесоводством, приобретает значение дарвиновского естественного отбора. Закон естественного отбора в лесу царит так же, как и во всем биологическом мире.
Установив факт наличности борьбы за существование, следует обратиться к изучению той закономерности, которая давно подмечена в ходе борьбы за существование в лесоводстве; вкратце я уже ее коснулся в начале нашего курса; сравнивая опытные таблицы разных пород, мы можем убедиться, что в насаждениях одной породы убыль числа стволов с возрастом идет быстрее, у других – медленнее.
Ниже я сопоставлю в одной таблице (табл. 44) соответствующие данные для трех пород: пихты, ели и сосны в различные возрасты.
Таблица 44
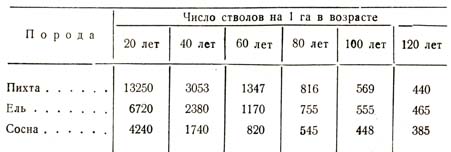
Из только что приведенной таблицы, как из всех таблиц, опубликованных до сего времени, мы усматриваем такую закономерность: процесс убыли, будучи неодинаковым у разных пород, находится в прямой зависимости от степени теневыносливости данного растительного вида; пихта теневыносливее ели, ель теневыносливее сосны, и в связи с этим большую густоту населения в любой возрастной момент мы наблюдаем у пихты, затем у ели и, наконец, у сосны. Чем теневыносливее порода, тем в каждый данный момент плотнее может быть население; количество выбывающих из строя, т. е. отмирание угнетенных, в сем случае идет меньшим темпом, чем у светолюбивых, потому что заглушённые деревца долго в состоянии влачить жизнь при недостатке света. Влияние указанной биологической особенности можно характеризовать следующими данными: в 50-летнем насаждении средняя площадь, занимаемая каждым индивидуумом, равняется:
у сосны........7,3 м2 = 100
ели .......6,4 = 87
бука .......5,8 , = 79
пихты........4,6 , = 63
то есть теневыносливые породы требуют 50 – 75% того пространства, которое необходимо для светолюбивых.
Шуберг показал, что если число стволов бука в любом возрасте принять за 100, то число стволов у других пород выразится следующими величинами [1 Большие исследования по развитию и строению леса в СССР были произведены проф. А. В. Тюриным и рядом других исследователей.] (табл. 45).
Таблица 45
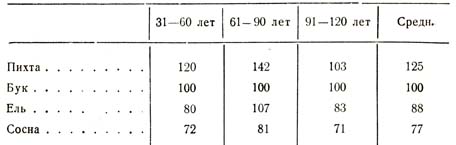
Нужно еще добавить, что у светолюбивых пород раньше наступает сомкнутость и, помимо более быстрого темпа, раньше начинается выдел угнетенных классов.
Такие примеры можно было бы увеличить во Много раз, но ввиду недостатка места ограничусь приведенными.
Вторая закономерность ясно видна из следующих сопоставлений (табл. 46)
Таблица 46
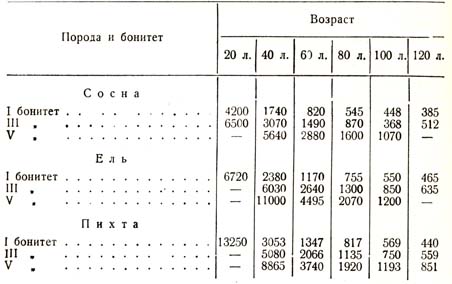
И сколько бы мы таких данных «и приводили, все они убеждают нас в том, что чем лучше условия почвы и грунта, тем энергичнее идет борьба за существование, тем меньшее число деревец или меньшую плотность населения мы наблюдаем в каждый возрастный момент, причем это справедливо по отношению к любой породе в любом климате. Это, казалось бы, на первый взгляд странное явление при ближайшем рассмотрении представляется совершенно естественным: чем лучше почвенная среда, тем богаче развивается каждая отдельная особь, тем раньше наступает сомкнутость, раньше деревья почувствуют взаимное стеснение друг другом и раньше начнется выделение угнетенных классов и борьба между ними.
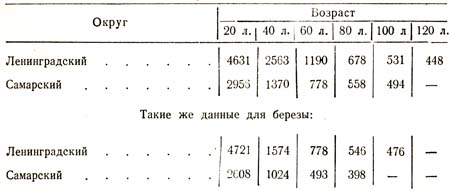
В таком же положении находится вопрос и о влиянии климата на ход борьбы за существование: чем благоприятнее климат данной породы, тем большей производительностью характеризуется ее насаждение, но тем и раньше начинается и быстрее происходит борьба за существование. Для иллюстрации приведу следующие примеры из наблюдений над сосною русского исследователя Варгаса-де-Бедемара в Ленинградском и Самарском округах (табл. 47).
Таблица 47
В таком же роде и влияние высоты над уровнем моря. Шуберг, разделив всю изученную им в Бадене область буковых лесов на три пояса – 1) до высоты 400 м над уровнем моря, 2) от 401 до 800 м и 3) от 801 до 1200 м, – получил среднее число дерев на 1 га для возраста от 31 до 120 лет, II бонитета, в первом случае (для первого пояса) 1291, для второго пояса 1510, для третьего пояса 2800, или, принимая первое число за 100, получим соотношение между числом стволов в трех поясах, равное 100: 126:244.
Следующим моментом, определяющим закономерный ход борьбы за существование, является возраст насаждения. Оказывается, что в сосновых насаждениях I бонитета Ленинградского окр., по данным Варгаса-де-Бедемара, выделяется за 60 лет, в возрасте от 20 до 80 лет – 3953 дерева, а в возрасте от 80 до 140 – только 281.
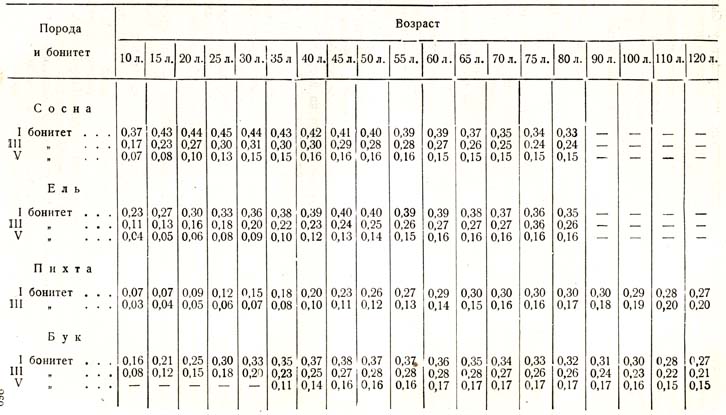
Мы видим с возрастом борьба затихает, и число дерев, переходящих из господствующих в угнетенные, все уменьшается [1 Отмечаем еще раз ошибочность и вредность мальтузианского объяснения процесса естественного изреживания насаждений. Ред.]. Если бы мы стали прослеживать быстроту этого процесса, то могли бы убедиться из всех табличных данных, что в общем более быстрая убыль приурочена к более молодому возрасту 20 – 40 – 60 лет, в зависимости от породы, климата и почвы; она медленнее происходит после указанного возраста. Дальнейший анализ показал бы нам связь этого явления с другим, именно: с наступлением большого периода роста, с кульминацией прироста в высоту. Не только исследования физиологов над ростом клеток, органов, тканей, но и лесоводственные наблюдения над приростом, хотя бы в форме тех опытных таблиц, о которых все время идет речь, превосходно показывают наличность закона большого периода роста. У так называемых быстрорастущих древесных пород этот период наступает весьма рано, у медленнорастущих – позже, у светолюбивых – раньше, чем у теневыносливых, на лучших почвах и в более благоприятном климате – раньше, чем в противоположных условиях, хотя абсолютная величина в первом случае будет больше, во втором меньше. Теперь приведу доказательства сказанного, а затем остановлюсь на значении этого явления (см. табл. 48).
С законом большого периода роста, с кульминацией прироста в высоту совпадает момент энергичного выделения угнетенных классов; таким образом, определенное явление, характеризующее общественную жизнь деревьев, в своем ходе подчиняется глубоко скрытым в протоплазме особенностям последней, так как закон большого периода роста не в своих колебаниях и изменениях внешних условий, а в самом существе своем глубоко коренится в неизвестных свойствах живого существа.
С ходом борьбы за существование тесно связан целый ряд явлений в жизни леса. Благодаря тесноте деревья, стоящие в лесу, вырастают выше и приобретают характерную форму – большую полнодревесность и малую сучковатость; деревья развиваются преимущественно в ствол и в меньшей степени развивается крона; благодаря борьбе за существование в лесу действует закон естественного отбора и происходит постепенное изреживание леса, а с этим процессом связаны многие явления: степень изменения обстановки под пологом, зарождение подлеска, появление подроста и т. д.
ВОЗОБНОВИТЕЛЬНЫЙ ПРОЦЕСС В ЛЕСУ
Возобновительная способность леса складывается, как было уже сказано, из трех крупных явлений, следующих во времени друг за другом: 1) плодоношение дерев, составляющих насаждение, 2) появление в ложе, приуготовленном лесом, самосева и 3) превращение самосева в подрост. Каждое из этих явлений зависит, помимо вмешательства человека, от причин трех категорий: 1) от биологических, внутренних свойств пород; 2) от внешних условий – климата, почвы, погоды и т. д. и 3) от социальных условий, создаваемых самим лесом. Покажем теперь зависимость каждого из вышеупомянутых трех процессов от указанных моментов и обратимся прежде всего к плодоношению деревьев.
Древесные растения в состоянии, как известно, приносить семена лишь с известного возраста возмужалости, который наступает в различное время у разных пород, а у одной и той же породы – в зависимости от климата, почвенных условий и того обстоятельства, растет ли дерево в сообществе с другими или свободно. Возраст возмужалости раньше наступает у быстрорастущих и светолюбивых пород, позже у медленнорастущих и теневыносливых, что находится, несомненно, в связи с ходом нарастания дерева в высоту и по объему, с кульминацией прироста, так как возраст возмужалости всегда совпадает с началом падения прироста. Раньше наступает возраст возмужалости у березы, тополей, ив, затем у лиственницы, сосны, дуба, ели, пихты и бука. Порода в оптимуме своих климатических условий плодоносит раньше, больше и чаще, чем в менее благоприятных климатических условиях. Возраст возмужалости наступает, как показали наблюдения лесоводов, раньше у деревьев, растущих на свободе, чем в насаждении, лет на 10 – 15, даже на 20.
Таблица 48
Текущий прирост в высоту в метрах
Производство семян требует значительного количества запасных веществ; наблюдения показали, например, что обильное плодоношение у бука вызвало уменьшение годичного прироста в 172 – 2 раза, а отложение запасных веществ на зиму в 3 – 4 раза; вот почему совершенно понятна должна быть зависимость между плодоношением и приведенными выше условиями, сводящимися, в конце концов, к условиям питания; вот почему также понятна должна быть связь между семеношением и ходом прироста. В насаждениях у деревьев позже наступает возмужалость благодаря худшему питанию и иному ходу роста, чем это свойственно свободно растущим деревьям, в частности благодаря меньшему развитию крон, более плохому развитию корневой системы, меньшему доступу света и тепла к кронам, более холодной и более сухой почве. Указанные обстоятельства, создаваемые насаждением, влияют не только на возраст возмужалости, повышая последний, но и на общее количество семян, которое дает та или иная порода. Исследования В. Д. Огиезского показали, что выросшие в насаждении, а потом выставленные на простор сосновые деревья, так называемые семенники, приносят в 17 – 20 раз больше семян, чем одномерные о ними деревья насаждения (рис. 76); опушечные же экземпляры, по данным того; же автора занимают среднее место, что и само: собою понятно, так как одной половиной они обращены к насаждению и, следовательно, принуждены испытывать все условия конкуренции, другой половиной, наоборот, – к простору, где нет соперничества других деревьев, где больше света и тепла, где их корням предоставлено больше места в почве, где крона их лучше развита, где, одним словом, условия питания лучше. Исследования В. Д. Огиевского показали, что у семенников большее количество мелких ветвей в кронах: на единицу площади сечения ствола приходится у них 100 – 125 кг, тогда как для дерев в насаждении 70 – 100 кг; мало того, каждый килограмм таких мелких веток отличается большей семеннон производительностью па просторе, чем в насаждении: в первом случае ветви несут от 5 до 12 шишек, во втором – от 1 до 3. Но не все деревья, образующие насаждение, принимают одинаковое участие в плодоношении; как показали исследования проф. А. Н. Соболева и А. В. Фомичева, в семенной год ели плодоносят только – I, II и отчасти III классы по Крафту, так как ничтожный урожай IV класса не может быть принят во вни-
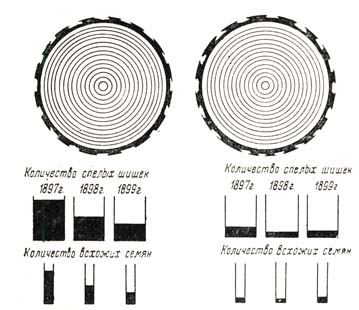
Рис. 76. Плодоношение сосны в Собинском лесничестве, Черниговский окр. Данные В. Д. Огиевского
мание, а V и совсем бесплоден. Ниже следует сводная таблица названных авторов (табл. 49).
Таблица 49

Из нее авторы делают следующие выводы:
1) 98% всего плодоношения в еловом насаждении приходится на долю господствующих в насаждениях деревьев, т. е. первых трех классов; 2% дают деревья IV класса, а V класс совершенно не участвует в плодоношении;
2) сравнивая между собою по участию в плодоношении первые три класса, видим, что первое место (45%) принадлежит II классу, второе (29%) – III классу и третье (24%) – I классу; 3) указанная последовательность зависит не столько от числа дерев каждого класса, сколько от степени плодоношения каждого дерева насаждения; именно, если мы разделим урожайность каждого класса на соответственное число деревьев, то получим:
для I класса..........24 : 8 = 3
II .........45:27 = 2
III .........29:28 = 1
IV..........2:17=1/8
Эти величины служат как бы показателем относительной урожайности дерева в каждом классе; мы видели, что дерево I класса приносит в 3 раза более, а дерево II класса в 2 раза более семян, чем дерево III класса, тогда как дерево IV класса – в 8 раз менее дерева III класса и в 24 раза – дерева I класса.
Как мы уже упоминали выше, эти различия находятся в зависимости от различных условий питания, в какие поставлены деревья разных классов.
Детальное исследование урожайности семян 1904 г. в одном еловом насаждении Охтенской дачи Ленинградского округа привело А. В. Фомичева к следующим выводам:
1) «если урожайность одного дерева I класса принять равной 100%, то урожайность деревьев остальных классов выразится следующими цифрами: И – 53,4%, III – 33,5%, IV – менее 0,2% и у – 0%»;
2) «несмотря на свою значительно большую семейную производительность, деревья I класса Крафта все же не играют роли в плодоношении всего исследованного насаждения и уступают свое место следующим двум классам. Это происходит потому, что число деревьев I класса очень незначительно (0,7%) и главный полог насаждения состоит из деревьев II (32,1%) и III (34,3%) классов Крафта. Оказывается, что 97,4% по весу от всего урожая семян в исследованном насаждении приходится на долю II и III класса, 2,5% – я а долю I класса и лишь около 0,1% на долю IV класса».
Задавшись вопросом, какая часть кроны деревьев в насаждении обладает наибольшей семенной производительностью, А. В. Фомичев получил весьма любопытные данные о величине урожайности различных двухметровых секций, на которые он разделил всю крону и плодоношение которых он изучал отдельно. Оказывается, что только дерево I класса, имеющее почти совершенно свободную и равномерно развитую крону, плодоносило равномерно на протяжении 8 м длины кроны, причем, однако, на нижних 2 м плодоношение было уже незначительное; первая или верхняя секция, находившаяся в особенно благоприятных условиях для плодоношения, дала 43,7% от всего урожая семян; 1вторая секция, находившаяся в условиях, одинаковых с первыми секциями крон деревьев II класса и составлявшая вместе с ним главный полог насаждения, дала 22,8% всего плодоношения, третья секция – 25,5% и, наконец, четвертая, наиболее затененная – всего лишь 8%. У дерев II и III классов, по исследованиям того же автора, плодоносит главным образом первая секция, дающая от 93 до 96% всего урожая и оставляющая на долю второй от 4 до 7%, лишь изредка больше (14%). Еще более сильно выражено отрицательное влияние на плодоношение угнетения кроны на деревьях IV класса, где шишки находились лишь в самых верхних частях первой секции, на второй – их совершенно не было. Итак, чем сильнее стеснена крона, тем меньшая часть ее способна плодоносить. По вычислениям
A. В. Фомичева, для первой секции крон в среднем на 1 кг живого веса кроны приходится около 12,7 г семян, для второй секции – 1,22 г, а для третьей – 0,70 г. Мы видим данные, до некоторой степени аналогичные тем, какие были получены B. Д. Огиевским, сравнившим производительность кроны свободно стоящих деревьев с теми, которые растут в насаждении.
Итак, мы видим, что произрастание деревьев в сообществе друг с другом изменяет следующие элементы плодоношения древесных растений: во-первых, увеличивает возраст возмужалости, во-вторых, уменьшает плодоношение дерева в насаждении по сравнению со свободно растущим, в-третьих, создает большое разнообразие в семеношении деревьев одного насаждения, но разных классов Крафта, и, в-четвертых, влечет за собою оставление потомства только господствующими, лучше развитыми, победившими в борьбе за существование деревьями. Совместная жизнь деревьев в насаждении влияет на плодоношение еще водном направлении, именно усиливая так называемую периодичность в наступлении семенных годов. «Несмотря на наступление возраста естественной зрелости, деревья приносят семена не каждый год: в некоторые годы, так называемые семенные, у дерев данной породы замечается особенно сильное плодоношение». В промежутке между полными семенными годами деревья также могут приносить семена, но только в меньшем количестве, и тогда различают половинные, четвертные и прочие семенные годы наряду с годами полного неурожая семян. Величина промежутка времени между семенными годами так же, как и другие элементы, характеризующие в своей совокупности плодоношение древесных растений, зависит как от породы, так и от тех условий климата, погоды, почвы, в какие поставлены насаждения. Если для плодоношения вообще необходима большая затрата со стороны растения энергии и питательных веществ, то в годы обильного урожая, или в семенные годы, это обстоятельство должно иметь особенное значение. Ввиду сказанного нет ничего удивительного, что частые семенные годы приурочены к благоприятным условиям роста в отношении климата, погоды и почвы: дуб, например, в Придунайских низменностях плодоносит почти ежегодно, в более теплых частях Германии – через 5 – 5 лет, у нас в Воронежском окр., по наблюдениям Корнаковского, в среднем через 7 лет, а в более холодном поясе, в Тульских засеках, например, кажется через 10 лет.
По некоторым данным средняя урожайность за 20 лет для отдельных пород колеблется от 15 до 45% от полного урожая; в порядке же последовательного уменьшения этой величины все породы распределяются следующим образом:
Береза.............. 45%
Граб ............. 42%
Ольха.............. 40%
Сосна.............. 38%
Ель.............. 37%
Пихта европейская........... 34%
Ясень.............. 33%
Дуб............... 17%
Бук............... 16%
Проф. А. Н. Соболев назвал эти цифры коэффициентами плодоношения. На основании их все исследованные породы можно разбить на три группы: в первую войдут наиболее часто плодоносящие, т. е. дающие каждые два года количество семян, почти равное полному урожаю, т. е. 100%, – сюда относятся береза и граб; ко второй группе относятся породы, дающие в три года столько семян, сколько бывает в один урожайный, – это ольха, сосна, ель, пихта и ясень; наконец, в третью группу входят породы, наименее богатые семенами, – дуб и бук, у которых лишь в 6 лет получается количество семян, равное полному урожаю.
По другим данным средняя урожайность для 10-летнего промежутка времени выражается следующими величинами.
Средняя урожайность семян в сотых долях полного урожая:
Бук – 0,25
Дуб – 0,29
Лиственница – 0,31
Ильм – 0,38
Сосна обыкновенная – 0,39
Пихта европейская – 0,39
Клен остролистный – 0,44
Ель – 0,45
Ясень – 0,46
Ольха черна – 0,50я
Береза – 0,54
Граб – 0,54
Здесь слабее плодоносят бук и дуб, а всего сильнее береза и граб.
У нас, по наблюдениям проф. Турского, в окрестностях Москвы, урожайными годами для сосны были:
1882..... обильный
1883..... средний
1884 .... обильный
1885..... средний
1886 .... слабый
1887..... средний
1888 обильный
1889 средний
1890 слабый
1891 обильный
1892 обильный
По данным того же автора, урожаи еловых семян в той же местности наблюдались в 1877, 1883, 1885 и 1890 гг.
Плодоношение насаждений до недавнего времени изучалось чисто глазомерным путем. Организованные германскими лесными опытными станциями наблюдения над урожаем лесных пород носили анкетный характер, причем лесничие должны были отмечать степень урожая (полный, средний, слабый и неурожайный) глазомерно, не отмечая еще при этом сопутствующих условий. Для перевода этих данных в цифровые данные принят полный урожай за 100, средний за 50, слабый за 25 и неурожай за 0 и, пользуясь обычной формулой для вывода средних величин, дал те цифровые выражения для урожайности разных пород, которые мы привели выше. То же самое сделал и Вимменауер, только он принял полный урожай за 1, средний за 2/3, слабый 7з, а неурожай также приравнял 0. Лишь в 1881 г. появилась работа Кинитца, который точно определил количество шишек и семян в одном сосновом насаждении, но так как работа эта стоит совершенно особняком, то совершенно справедливо проф. А. Н. Соболев видит начало эры изучения плодоношения в работах В. Д. Огиевского, который с 1895 г. начал систематически работать над вопросами плодоношения, применив объективный метод исследования.
Изучив,. как влияет выставление сосны на ее плодоношение (некоторые из результатов этой работы мы приводили выше), В. Д. Огиевский обратился затем к определению того, как часты и как велики бывают урожаи сосновых семян и от каких условий зависит размер урожая. Для определения урожайности применялся точный учет шишек по модельным деревьям, которые выбирают из экземпляров III класса по Крафту; определялось число однолетних и двухлетних шишек; всего с 1895 г. было исследовано 550 деревьев в сосновых насаждениях в Черниговской и Орловской губерниях. Выводы были получены следующие: 1) урожайность семенников превосходит урожайность деревьев в насаждении в 4 – 6 раз (см. рис. 76); 2) средняя урожайность выражается следующей цифрой – 900 двухлетних шишек на Дереве III класса толщиною 53 см; 3) особенно урожайным был 1897 г., самый неурожайный 1900, значительные урожаи были в 1901 и 1902 гг. и, наконец, 4) «разница в урожайности отдельных годов зависит от количества осадков, выпадающих осенью и зимою перед образованием однолетних шишек: чем больше осадков выпадает с октября по февраль, тем более образуется в наступающем году однолетних шишек и тем более будет трехлетних шишек через два года».
Последний вывод встретил в лесоводственной литературе верные критические замечания, которые заставляют сомневаться в правильности вывода, указывая на недостаток метода. Автор сопоставляет урожаи шишек за известные годы и сопоставляет их с одним каким-либо климатическим фактором, причем количеством осадков в определенный период времени исследователю удалось выяснить определенное соотношение, тогда как ни с количеством света, ни с температурными условиями и количеством урожая связи обнаружить не удалось. Развитие растений зависит, как известно, от света, теплоты и влаги, а раз это так, справедливо замечает В. И. Иванов, то метод сопоставления данных урожая с влияющими на него факторами должен быть совершенно иной. «Метод, выбранный автором, был бы верен лишь в том случае, если бы на плодоношение сосны влиял только один какой-либо из приведенных факторов: или свет, или теплота, или влага; если известное явление зависит от целого ряда причин, то влияние какой-либо одной из них может быть выяснено при сопоставлении таких явлений, которые были вызваны причинами, действующими на сравниваемые явления в одинаковых количествах, за исключением одной какой-либо причины; тогда можно предполагать, что различие сравниваемых явлений было обусловлено причиной, действующей в различных количествах». В данном случае, по мнению В. И. Иванова, для выяснения влияния осадков на урожай сосновых шишек автор должен был бы сравнивать урожай шишек только за те годы, за которые и свет, и теплота действовали бы в одинаковых количествах, количество же осадков было разное, тогда только и можно было бы говорить о влиянии осадков на урожай сосновых шишек.
Последний вывод В. Д. Огиевского вызывает сомнение еще и потому, что, пользуясь таким же методом, проф. Шварц пришел к совершенно противоположному выводу, показав, что количество осадков с мая по июль имеет решающее влияние на ход роста сосны в толщину. Одно из трех: или прав один, или прав другой, или, наконец, оба неправы, но без сомнения, что раз обильные осадки вызывают обильное плодоношение, то не могут они вызвать противоположные явления – обильный прирост; здесь, повидимому, причина скрывается не в осадках, а в более сложном сочетании метеорологических факторов, в одном соединении создающем благоприятные условия для обильного плодоношения, в другом своем соединении и иной последовательности создающем, наоборот, благоприятные условия для усиленного прироста.6
В. Д. Огиевский применил объективный метод для учета плодоношения, но выбрал не вполне удачно в качестве представителя насаждения дерево III класса по Крафту. Исследования проф. А. Н. Соболева и А. В. Фомичева в методологическом отношении подвигают вопрос вперед, так как базируются на учете семян с модельных деревьев всех классов по Крафту, причем для наиболее многочисленных классов, как II и III, которые объединяют деревья с весьма различными по развитию кронами, названные исследователи раздробили указанные классы еще на несколько частей, так что в конечном счете они получали от 7 до 10 групп деревьев, вершины которых, по глазомерному определению, были более или менее однородны. Расчленив, таким образом, все деревья какой-нибудь пробы на 7 – 10 групп однородных в семенном отношении, авторы брали, как они выражаются, семенные модели в количестве 10% от общего числа деревьев. Какой же запас семян находится в урожае еловых насаждений, которые изучали названные авторы? Оказывается, что в одном случае на 1 га насаждение дало 13 000 кг, в другом 80 800, в третьем же случае из Охтенской дачи, при взятии насаждения 75 лет, 1,0 полноты, I бонитета и 1-й добротности, – 90 000 кг, это количество семян заключалось в 121 400 шишках; число же семян, составлявших урожай, достигло 21000 000, из которых всхожих оказалось около 14 500 000.
Совсем другой метод для изучения плодоношения был предложен проф. М. М. Орловым и, независимо от него, лесничим Замараевым. Исходя из идеи дождемеров, названные исследователи предложили устраивать особые плоские, открытые сверху деревянные ящики квадратной формы. В семеномерах, высотою в 12 см, устроенных по системе проф. М. М. Орлова, в дне ящика высверливаются по три круглых отверстия, которые прикрываются жестяными ситами, служащими для дренирования этих аппаратов во время дождливой погоды; сверху ящики закрываются проволочной сеткой. Замараев устраивает ящики с боковыми стенками высотою в 14 см, с поверхностью 0,5 м2, с отверстиями для стока, с защитной сеткой сверху. Последнее время и В. Д. Огиевский счел удобным пользоваться такими семеномерами, но устроенными по другому типу, притом из жести, в виде особых воронок. Оттянутая часть воронки покоится на земле и, служа для сбора упавших семян, тем самым предохраняет их от склевывания птицами, сноса ветром и т. п.
Наблюдений с помощью таких семеномеров опубликовано пока немного, и они притом носят предварительный характер. Замараевым были поставлены четыре семеномера среди чистого 100 – 120-летнего елового насаждения: два под пологом леса, третий на опушке леса, а четвертый на сплошной лесосеке, в расстоянии 21 м от одной стены леса и 42 м – от другой. Опавшие семена выбирались каждый день, причем велись некоторые метеорологические наблюдения; наблюдения были начаты 24 марта (1905 г.), и в период времени по 9 мая было в семеномерах насчитано следующее количество семян: в № Л – 378, в N° 2, – 323, или в среднем под пологом насаждения на 0,5 м2 350 шт.; в № 3 (на опушке) – 272, а в № 4 (на лесосеке) только 38. По месяцам опавшие семена распределялись так (табл. 50):
Таблица 50
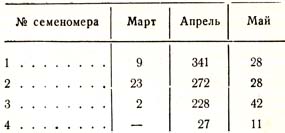
В семеномер на опушке, как мы видим, попало больше половины того количества, которое попало в приемники под пологом; это хорошо вяжется с ранее указанным фактором о большем плодоношении энергичных деревьев. Наибольшее количество семян выпало с 16 по 19 апреля, когда температура в 2 часа дня поднималась до 21 – 22°; наибольшее количество семян выпало при южном, юго-западном и юго-восточном ветрах. Принимая в 1 кг еловых семян 166 000 штук семян, Замараев находит, что урожай гектара исследованного насаждения равен 47,1 кг, тогда как принято высевать при сплошном посеве на 1 га от 7 до 13 кг. Мы можем сказать, что природа сеет полными руками.
Другое исследование опубликовано проф. И. И. Сурожем и относится к сосновым насаждениям дачи Руда (б. Люблинской губ.). Всего был установлен 21 ящик, каждый в 1 м2. Из этого исследования явствует, что указанная величина ящиков вполне достаточная, что семеномеры, поставленные в однородные условия, дают сравнимые результаты. В течение 4 лет – с 1902 по 1905 гг. – опало семян на 1 м2 для всех ящиков 292, при колебании по отдельным приемникам от 51 до 537. Оказывается, что в насаждении I класса возраста опало 51 семя, II класса – 268, IV класса – 442, VII класса – 496. Если количество семян, опавших во все ящики за 1905 г., принять за 1, говорит проф. Сурож, то для 1904 г. оно выразится 19, для 1903 г.- – 3, для 1902 г. – 2 и для (1901 г. – 2. Таким образом, продолжает тог же автор, за 5 лет учета имел место лишь так называемый один семенной год с плодоношением в общем в 6 – 18 раз большим сравнительно с другими годами данного пятилетия. Опад семян начался около половины апреля, продолжался в мае, захватывая даже июнь месяц, но массовый вылет, в количестве более 50% всего годового учета, обыкновенно, по Сурожу, приурочивается к сроку малой продолжительности – в 7 – 15 дней, причем время наступления его находится, по-видимому, в зависимости от высоты температуры воздуха. При расчете, что в семенной год в среднем на 1 м2 оказалось 292 семени, на гектар это составит около 2,9 млн. шт., или около 14 кг.
По этим, как мы видим, пока отрывочным, но весьма интересным данным мы можем судить, как будут чреваты последствиями наблюдения над плодоношением, изучаемым только что изложенным методом. Строго говоря, этим методом изучается не плодоношение насаждения, а то, как оно реализуется и закрепляется в почве, занятой лесом или находящейся по соседству под его влиянием.
При изучении возобновительного процесса надо иметь в виду нее последовательные стадии его, надо подвергнуть объективному наблюдению не только величину плодоношения самого насаждения, но и реализацию его или закрепление его почвой, приступив затем к такому же объективному учету количества появившегося самосева и следя далее за его развитием, отпадом, ростом и т. д.; следует продолжать наблюдение и тогда, когда самосев создает уже сомкнутые группы подроста. В плодоношении деревьев играет большую роль индивидуальность их, но тем не менее явления, характеризующие семеношение, так же закономерны, как и другие явления природы, – они так же могут е успехом изучаться с надеждой вскрыть причинные зависимости, как и другие явления в жизни леса.
Переходим теперь к исследованию тех условий, которые определяют прорастание семян или появление самосева. Для прорастания семян, как известно, необходимо определенное количество тепла и влаги; последний элемент обеспечен в лесу, так как самые верхние горизонты почвы, прикрытые мертвым покровом, будучи защищены от инсоляции, всегда в лесу влажнее, чем на открытом месте; мало того, влажность этих слоев в лесу характеризуется постоянством, тогда как поверхность открытой почвы, освещаемая лучами солнца, может, правда, быстро намокать, но так же быстро и высыхать и притом настолько, что лежащие на них или в них семена не в состоявши будут прорасти. Проф. В. Я. Добровлянский обращал внимание еще и на то, что влияние интенсивного освещения будет различно по отношению к семенам различных древесных пород. «Чем крупнее семена данной породы и чем длиннее для них период семенного покоя, тем труднее будет им воспользоваться нужным количеством быстро испаряющейся влаги; очень мелкие и быстро прорастающие семена, например березы ли сосны, будут иметь возможность прорасти даже и на совершенно открытой площади, где влага удерживается очень короткое время».
Что же касается тепла, то, как мы знаем уже, почва и атмосфера под насаждением получает его меньше, чем открытое пространство, но это обстоятельство выгодно для пород, чувствительных к заморозкам, боящихся или в стадии всходов, или даже в первые годы своей жизни солнцепека. Следующими благоприятными моментами для прорастания семян под пологом леса являются для одних, опадающих осенью или летом, – заделывающая деятельность той новой подстилки, которая создается после опадения листвы, для опадающих же ранней весной, еще по снегу, – рыхлость и постоянство снежной пелены, сквозь которую, если она не образует наста, темного цвета семена быстро и легко проникают до почвы.
Затенение и все другие условия, связанные с пологом, создают, как известно, своеобразный тенелюбивый живой покров в лесу, который не мешает ни прорастанию семян, ни дальнейшей жизни всходов. На открытых местах, где светолюбивые злаки образуют живую густую дернину, в конце концов создается целый густой войлок из отмерших ее частей, который, как показали наблюдения В. Д. Огиевского, препятствуют часто корням прорастающих семян достигать минерального слоя почвы; правда, так называемые зависшие всходы встречаются и в лесу, в, слое толстого мохового или мертвого покрова, но это случаи, так сказать, патологические. Под пологом насаждения всходы обычно не встречают столь сильную конкуренцию со стороны травяного или вообще живого покрова, как вне леса, где он действует, как соперник, и своими корневыми и своими надземными органами.
Неблагоприятными моментами для последующей жизни всходов в лесу являются следующие:
1. В одних случаях – в насаждениях сложных, густых, в особенности при участии теневыносливых пород в их составе – недостаток света, особенно чувствительный для пород светолюбивых.
2. В некоторых случаях решающую роль может сыграть недостаток влаги в. верхних горизонтах почвы, случай – на каком мы уже останавливались, когда речь шла о чистых, одноярусных светлых сосновых насаждениях в сухом климате и на сухих почвах, где, как было указано в своем месте, корневая система материнских насаждений сильно развивается в поверхностном направлении и тем самым усиливает конкуренцию для самосева. В этих так называемых сухих борах самосев сосны, появляясь в семенной год весною, часто сохраняется только в свежих низинах между холмами, отмирая уже к середине или к концу лета на буграх и склонах. Необходимо сочетание благоприятных условий испарения в течение нескольких лет подряд, чтобы в подобных положениях самосев сохранился; необходимо добавить еще, что в указанных условиях самосев не испытывает недостатка света или конкуренции травяного покрова, крайне редкого в таких борах; нет здесь также и вредного влияния кислого плотного, перегноя, так как почва почти не прикрыта редкой опавшей хвоей. Отсутствие каких-либо других реальных факторов, которыми можно было бы объяснить умирание самосева и приуроченность последнего явления к засушливому периоду, в связи с доказанной многочисленными определениями большой сухостью почвы, падающей ниже своей двойной гигроскопичности, в связи с противоположными явлениями, разыгрывающимися в свежих низинах, где самосев сохраняется, несмотря на травяной покров, – все это заставило меня в ряде работ высказать обоснованное положение, что в сухих борах самосев сосны может погибать не от недостатка света или других причин, а от недостатка влаги, несмотря на защиту материнского полога.
3. Может под пологом леса погибать самосев и от наличности плотного и мощного кислого перегноя, который мешает корням проникнуть до минерального, слоя почвы. В исследовании моего ученика А. Новака, произведенном в Буаулукском бору в сложном насаждении из сосны р лиственными породами {дуб, липа, береза, осина) и кустарниковым подлеском на пермских мергелях, указанный момент влияния мертвого покрова на количество самосева сказывается довольно ярко.
Здесь, в этих сложных насаждениях, подстилка достигает большой мощности и большого разнообразия как в этом отношении, так и в отношении рыхлости, колеблясь от 0,3 до 7,0 см. Путем заложения пробных площадок (в количестве 50 шт.) в различных условиях А. Новаку удалось показать следующую закономерную связь между мощностью подстилки, с одной стороны, и количеством самосева, с другой (табл. 51).
Таблица 51
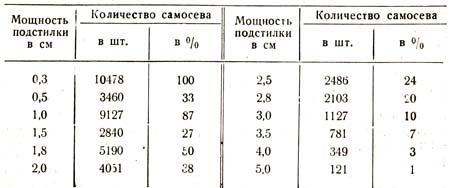
При мощности подстилки в 6 – 7 см самосева совершенно не было. В приведенных данных закономерность нарушают только вторая и четвертая группы сверху, давшие слишком преуменьшенный результат сравнительно с тем, который они по положению в ряду должны были бы дать. Однако исключения только подтверждают правило: подробные данные в основной таблице показывают, что, в то время как первая, третья и пятая группы обладают покровом рыхлым, без признаков плесени, вторая и четвертая пробные площадки имеют покров более плотный, с ясно выраженной плесенью, что и сыграло здесь решающую роль. Те же пробные площади были затем распределены на группы по степени плотности покрова; полученные данные свидетельствуют о закономерности и в этом направлении, как доказывает следующая таблица:
Рыхлый покров.........3462 шт. или 100%
Средней плотности покров.....1560 или 75%
Плотный покров..........406 или 12%
Степень благоприятности условий для прорастания семян и для жизни всходов в первые годы выражают в лесоводстве термином возобновительная спелость почвы. К указанным выше моментам, определяющим эту спелость, следует отнести еще качество гумуса, благоприятное для одной породы и действующее для другой как яд; но этот важный фактор так мало еще изучен, что пока останавливаться на нем не станем. Можно еще указать и на другие причины плохого обсеменения под пологом леса, Например на избыток влаги плохо дренированных мест или солонцеватость почвы в нашей лесостепи, но это – все обстоятельства, не вызванные самим насаждением, а связанные с определенным климатом, равнинностью места и другими- моментами внешней среды; и сами насаждения от них сильно страдают, страдает и их обновление.
Если мы будем присматриваться к распределению самосева под пологом насаждений, то мы в редких случаях найдем однородность в этом отношении; обыкновенно же самосев распределен неравномерно и большею частью группами, на первый взгляд, капризно распределенными по поверхности почвы. На самом деле, однако, здесь нет каприза, а хаотическое, на первый взгляд, распределение групп самосева и подроста есть только, отпечаток степени возобновительной спелости лесной почвы, неодинаковой в разных местах насаждения. Это сильно подчеркнуто многими лесоводами, которые, между прочим, и на этом основании требовали не равномерных обсеменительных рубок, а неравномерных. Что же касается причины того, почему спелость почвы под пологом леса неодинакова, то здесь повинны все почвообразующие факторы: и состав материнской породы, если он не совсем однороден, и рельеф поверхности, и, наконец, состояние насаждения, в особенности его большая или меньшая полнота. В сухих условиях самосев и подрост приурочен больше к потным и более влажным местам; в мало-дренированных местах – к более возвышенным; на болотистых – к кочкам, к валежнику и т. п.; при более или менее однородных почвенногрунтовых условиях – к так называемым окнам среди насаждений, т. е. небольшим прогалинкам.
По исследованиям моего ученика Л. Ф. Рудовица в Засурской даче, Пензенского окр., в сосновых насаждениях надлуговой террасы, на основании заложенных проб на количество самосева, последнего оказалось в среднем 2293 экземпляра на гектаре, во со значительными колебаниями – от 46 до 9688 шт. Подрост по большей части расположен в окнах, местах, освещенных сверху, образуя здесь очень густые группы: в месте, освещенном сверху, при прочих равных условиях – 4932 шт., на площадке, сверху затененной, – только 1806 шт. Влияние густоты насаждения на возобновление особенно хорошо иллюстрируется теми местами, где было искусственное изреживание, так называемая вырубка полумассы в целях вызвать обсеменение площади; здесь вместо обычного количества подроста, т. е. 2281 шт. в среднем, Рудовиц нашел 23 265 шт. на гектаре, причем там, где полнота приближалась к 0,7, количество падало до 12 191 шт. на гектаре. Новак в упомянутой работе показал, что в сложных сосновых насаждениях Бузулукского бора количество подроста сосны колеблется, в зависимости от полноты насаждения, следующим образом:
при полноте 0,9........583 шт. 18%
0,8.......1365 42%
0,7........1250 36%
0,6........3205 100%
0,5........2477 78%
0,4.......2472 , 76%
0,3........440 , 13%
0,2........286 8%
Перед нами два примера сосновых насаждений, имеющих ту общую черту, что количество самосева в каждом из них подчинено) одной и той же закономерности: максимум его совпадает с некоторым оптимумом полноты – с некоторой средней полнотой 0,5 – 0,6; вверх и вниз от этой гущины леса одинаково уменьшается количество самосева, хотя и по разным причинам.
Имея эту общую черту, оба указанных сосновых насаждения весьма резко отличаются друг от друга другой особенностью: в одном случае уменьшение полноты до оптимальной увеличивает количество самосева в 10 раз, в другом в 5 с небольшим; в одном случае мы получаем весьма большое количество, в другом, в конце концов, все же незначительное; насаждения первой группы отличаются, по нашему мнению, большой возобновительной упругостью, другие в этом отношении – мало упруги и потому, если даже могут дать такой же эффект, то не одним лишь изменением полноты, но и другими искусственными приемами, о которых здесь не место распространяться.