Опыт был организован так, что убыль в весе всегда могла быть приравниваема к потере влаги от испарения ее растением. Для большинства взятых пород опыт продолжался три вегетационных периода, и в нижеследующей таблице показано количество испарившейся влаги в килограммах (среднее на три вегетационных периода), отнесенное на 100 г сухого вещества листьев:
Ясень........... 85,6
Береза........... 81,4
Бук .......... 74,8
Граб........... 73,0
Вяз....... ... 66,2
Дуб .......... 54,6
Клен остролистный....... 53,0
Ель .......... 13,5
Сосна обыкновенная....... 9,4
Пихта........... 7,2
Сосна черная......... 6,7
Из этих данных мы можем видеть прежде всего, что хвойные испаряют гораздо меньше, чем лиственные, – вывод, который, как мы видели уже, был сделан Клаубрехтом. По вычислениям Гёнеля, испарение хвойных относится к испарению лиственных, как 1 : 10.
Исследования Гёнеля обнаружили еще следующие интересные факты:
1. Количество испаряющейся влаги меняется с интенсивностью поливки в прямом отношении, т. е. с увеличением поливки увеличивается и количество расходуемой растением влаги: например, ясень при минимуме поливки испаряет на 100 г сухого вещества листьев 55 кг, а при максимуме – 98; сосна в первом случае – 5, во втором – 12,1. Этот факт показывает, что испарение есть процесс не только физический, но и физиологический, который регулируется растением.
2. Количество испаряющейся влаги зависит от того, помещается ли растение в тени или на солнце, причем важно то обстоятельство, что когда у сосны и пихты в тени уменьшается транспирация, у бука и граба, наоборот, она увеличивается, как доказывают данные табл. 22.
Таблица 22
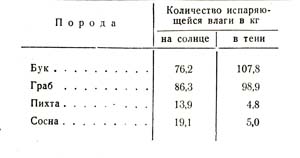
Это находится в связи с деятельностью устьиц, которые, оставаясь в тени у лиственных пород открытыми, заставляют их больше (расходовать в этих условиях влаги.
Исследование Гёнеля имеют, конечно, тоже свои недостатки: в сосудах помещалось 3 – 5 кг земли, что для 5 – 7-летних деревец надо признать количеством недостаточным; герметическая укупорка обусловливала недостаток кислорода для дыхания корней, чем, вероятно, и объясняется большая смертность среди опытных растений; по всей вероятности, кроме того, и температура внутреннего помещения, несмотря на защиту наружным сосудом, была более высока, чем это нужно. Кроме того, Генель вычислял испарившуюся воду только на часть того вещества, которое приросло за вегетационный период; он, как мы видели, вычислял на вес листьев, не принимая во внимание прироста ствола и корней. Притом у хвойных годичный прирост хвои составляет только1/3 – 1/5 часть всей хвои, и потому, по справедливому замечанию проф. Л. А. Иванова, для сравнения испарения хвойных с лиственными числа для первых нужно было увеличить в 3 – 4 раза.
Вот тот материал объективных исследований, которым располагает лесоводство для освещения вопроса об отношении Древесных пород к влаге. Попробуем несколько разобраться в этом малоизученном, но чрезвычайно важном вопросе.
Прежде всего, как учит физиология растений, надо отличать скорость, или интенсивность, испарения от экономности испарения и продуктивности его. Первая определяется исчислением количества испарившейся влаги на единицу поверхности растения. Перечислением того же количества на содержащуюся в растении воду дается понятие об экономности испарения; один из исследователей определил, например, что экономность испарения сосны вдвое больше чем у ели, между тем как по интенсивности испарения эти две породы очень близки друг к другу. Тот же исследователь показал, что пятилетние деревца сосны и дуба потребляют за сутки только часть содержащейся в них воды, именно около 75%, тогда как ель и особенно бук испаряют за сутки в 1 1/2 – 2 раза больше того количества, которое содержится в них; иначе говоря, у этих пород в течение суток запас воды должен возобновляться несколько раз. Если делать перечисление на вес сухого вещества, то мы получаем понятие о продуктивности испарения, а не об интенсивности его, как полагали раньше.
Совокупность этих понятий может дать представление о потребности древесных пород в отношении влаги. Но кроме потребности следует отличать как и в отношении зольных веществ, требовательность пород к влаге, т. е. отношение их к условиям влажности определенной среды и уменье взять необходимое количество влаги в тех или иных условиях. Само собой разумеется, что потребность во влаге всегда будет одним из элементов, из которых слагается требовательность, но большая даже потребность во влаге еще не может быть синонимом большей требовательности; например, береза, по исследованиям некоторых авторов, потребляет большое количество воды, а между тем она в состоянии произрастать в довольно сухих условиях. Здесь можно отметить полную аналогию с потребностью и требовательностью древесных пород в отношении зольных веществ и общность некоторых моментов в организации растений, которыми можно объяснить как большую, так и малую требовательность. Величина поверхности корневой системы и величина ее деятельной части, а также глубина распространения и степень пластичности ее будут играть такую же существенную роль в снабжении влагой, как и в снабжении зольными элементами. Исследования ряда ученых в этом направлении, упомянутые в своем месте, имеют такое же значение и здесь при расценке требовательности на влагу сосны, ели и пихты. Незначительно различаясь в отношении потребности, эти три породы существенно уже отличаются друг от друга по пластичности, глубине распространения и величине поверхности корневой системы; оттого сосна может мириться с почвами более сухими, а ель и пихта требуют более влажных грунтов. Но, кроме устройства воспринимающей влагу системы, для освещения вопроса о различной требовательности пород к влаге необходимо еще обратить внимание на организацию листовой поверхности. Здесь имеют значение величина поверхности, форма листьев, толщина эпидермиса или кутикулы, волосяные покровы, восковой налет, способность менять положение и величина устьиц, величина листьев, время опадения, быть может, эфирные масла и т. д. Должно иметь также значение в этом вопросе соотношение между корневой поверхностью и листовой, число проводящих путей в древесине, а также величина растений, но все это в применении к древесным породам, так мало еще изученным, что можно лишь привести отдельные примеры: у сосны, например, устьица погружены ниже уровня листовой поверхности, игольчатая форма листьев хвойных уменьшает листовую поверхность по сравнению с породами, имеющими плоские листья; кожистость листьев наблюдается у хвойных, у падуба; профильное положение листьев наблюдается у можжевельников, а способность принимать таковое – у представителей семейства мотыльковых и т. д.
В отношении требовательности на влагу ботаники давно различали два биологических типа растений, именно: ксерофитов, или сухолюбов, и гидрофитов, или влаголюбов. Иногда образуют еще третью группу – мезофитов, находящуюся посредине между двумя крайними. Прежнее представление о сухолюбивом растении как о таком, которое способно населять сухие местоположения, является односторонним; в природе имеет гораздо большее значение не одна физическая сухость, а физиологическая – понятие гораздо более широкое. Известно, что не все количество влаги, находящейся в почве, доступно растениям. Некоторое количество влаги, разное в разных случаях, является недоступным, как бы мертвым запасом для растений. В природе могут быть такие условия, что влаги в почве в физическом смысле будет вполне достаточно, но эта влага будет мало доступна ее потребителям или в силу своей соленосности, или обилия в ней гумусовых кислот, или, наконец, в силу ее низкой температуры; так бывает на солонцах, на болотах и вблизи горизонтов вечной мерзлоты. Но этого недостаточно для полного представления о физиологически сухих местах: важно обратить внимание не только на количество влаги и даже на количество доступной растению влаги, но и на у ел о в и я испарения ее. Одна местность с определенной влажностью почвы может находиться в спокойном от ветра месте, другая может быть расположена в условиях полного доступа ветра, например, но побережьям марей, на некоторых экспозициях IB шорных странах; с этой точки зрения важен также северный или южный склон и т. д. Вот почему понимаются под физиологически сухими местами, во-первых, такие, в которых затрудняется приход или поглощение влаги, во-вторых, такие, где недостаток влаги может обнаружиться благодаря чрезмерному испарению, и, в-третьих, такие, где комбинируются два Указанных условия. Поглощение влаги может быть затруднено как недостатком ее в почве или сухостью ее, так и наличностью тех элементов, о которых было выше упомянуто и которые могут делать почвы с достаточным количеством влаги физиологически сухими. Чрезмерные испарения могут быть вызваны: сухостью воздуха, высокой температурой его, сильным движением его, большим доступом света. Те растения, которые в своей организации приспособлены к одному из трех указанных условий, и будут называться ксерофитами, или сухолюбами. Большинство древесных пород нашей флоры принадлежит к влаголюбам или к средней категории – мезофитам, но некоторые, прежде всего сосна обыкновенная и черная, а также горная, тамариксы, терн, лох, некоторые ивы, можно отнести к сухолюбам. Можно для пород средней или повышенной требовательности на влагу устанавливать особую категорию – тропофитов, т. е. растений, которые в физиологически сухое время года, именно зимою, являются ксерофитам», а в физиологически более влажную пору года – мезофитами или гидрофитами. Действительно, зимою породы, лишаясь своих листьев, являются организмами, хорошо защищенными от испарения коркой, пробковыми образованиями, почечными чешуями и т. п.
Остановимся теперь на нескольких примерах прежде всего на самом ярком ксерофите из древесных пород – на сосне обыкновенной. Малая потребность в воде, доказанная опытами и Гёнеля и его предшественников, малая поверхность листовых органов в связи с самой формой хвои и ее анатомическим устройством – толстая кутикула, окруженная устьицами, – все это создает благоприятные условия для испарения. С другой стороны, большая корневая поверхность, могущая благодаря своей пластичности хорошо приспособляться к тем или иным условиям влажности, в свою очередь создает благоприятные условия для прихода влаги. Таким образом сосна является как бы хвойным ксерофитом, или сухолюбом, по двум основаниям. Исследования показали, что там, где грунтовые воды очень далеко удалены от поверхности, сосна развивает небольшой стержневой корень, но сильно развитую Поверхностно стелющуюся систему, приуроченную как бы к использованию атмосферной влаги. Там же, где почвы влажнее, и в особенности если грунтовые воды не далеки, развивается более глубокий стержневой корень, часто с характерными окончаниями1 в виде мелких боковых, утолщенных на конце, корней {см. рис. 64), сосущих воду из капиллярного уровня нижележащей грунтовой воды. Сосна, как известно, встречается на моховых болотах, где хотя ее рост и плохой, но все же она может мириться с избытком физической влаги, с недостатком дыхания для корней, с обилием гумусовых кислот, делающих подобные места физиологически сухими.
Ель имеет несколько большую потребность во влаге, но главное – большую листовую поверхность, так как хвоя ее держится на дереве 5 – 7 лет (а у сосны 2 – 3 года) и имеет не столь резко выраженную ксерофитную организацию; корневая поверхность ее выше, она гораздо менее пластична и более поверхностна. В силу последнего обстоятельства, она должна нужное ей количество влаги взять из почвы и грунта меньшего объема, чем тот, который находится в распоряжении сосны. Потому для ели естественно представляется два выхода: или этот грунт должен отличаться большей влажностью, или условия испарения должны оказаться пониженными, благодаря, например, наличию влажного климата.
ОТНОШЕНИЕ ДРЕВЕСНЫХ ПОРОД К ТЕПЛУ
Потребность в тепле у разных древесных пород различна. И в этом отношении можно также различить разные биологические типы, как их давно и различают в ботанической географии. Но тогда как для распознавания теневыносливых и светолюбивых пород, сухолюбов или влаголюбов существуют объективные признаки, для различения разной степени теплолюбивости, как справедливо замечают многие, нет никаких данных Почечные чешуи, различные корковые образования, кутикула и т. п., будучи, правда, дурными проводниками тепла, не говорят еще об отношении к теплу, а представляют собой скорее приспособление против излишнего испарения. Не имея ни в наружном габитусе, ни в каких-либо признаках внутреннего строения указателей для отнесения породы к теплолюбам, к холодолюбам или к какой-нибудь средней группе, приходится довольствоваться одним путем, именно исходить из тепловых условий ее области роста. Зная область географического распространения породы, охарактеризовав так или иначе тепловые условия в пределах такой области, мы в состоянии составить даже шкалу теплолюбивости, как, например, следующая, принадлежащая одному из лесоводов: ильмовые, каштан, дуб, пихта, бук, липа, сосна, явор, береза, ясень, ольха, кедр, горная сосна, лиственница. В этой группировке едва ли правильно положение сосны, березы и ясеня.
Классификация пород в отношении требовательности их к теплу одним из лесоводов давалась по зонам. Прежде всего устанавливались типические породы в отношении климата, к каковым причисляет благородный, или съедобный, каштан (Castanea), бук (Fagus), ель (Picea), лиственницу (Larix). Соответственно этим типичным породам он расчленяет Европу, за исключением субтропического пояса, на такие тепловые зоны, как Castanetuni, Fagetum, Piceetum и Laricetum, давая каждой из этих областей краткую климатическую характеристику и перечень ей свойственных пород.
В общем же породы можно разместить в отношении их требовательности к теплу в следующем порядке: каштан, дуб, ясень, ильмовые, граб, приморская сосна, австрийская сосна, обыкновенная сосна, рябина, ольха, береза, пихта, ель, кедр и лиственница.
Такая неустойчивость метода чрезвычайно досадна ввиду первенствующей роли такого фактора, как тепло. В каждой тепловой зоне мы найдем своих светолюбов и тенелюбов, своих ксерофитов и влаголюбов, свой ряд требовательных к составу почв и нетребовательных растений; но теплолюбивые растения не в состоянии произрастать нормально, по крайней мере, в климате с недостаточным количеством тепла и обратно. Перенесение породы в более теплый или более холодный климат, как увидим сейчас, отражается на всех других лесоводственных свойствах породы. Есть еще обстоятельства, на которые нужно обратить внимание: тепловая энергия не отделима от световой, принимая же во внимание, что всякий полог леса отнимает часть тепла от почвы, что под пологом всегда холоднее, чем вне его, мы во многих случаях на границах распространения какой-нибудь породы не всегда в состоянии точно сказать, отчего преждевременно отмирает подрост под материнским пологом: от недостатка света или от недостатка тепла.
Для характеристики породы в отношении требовательности к теплу необходимо знать не только общую сумму тепла, но и продолжительность вегетационного периода, в течение которого реализуется это общее количество, а также отношение к крайностям температуры. Раньше остановимся на отношении пород к крайностям температуры, а затем уже на общем значении теплового фактора в жизни древесных растений. Необходимо различать ранние и поздние утренники, или заморозки. К поздним, или весенним, чувствительны следующие породы: пихта, ель, бук, дуб, реже ясень и клен; нечувствительными к таким заморозкам считаются береза, лиственница, сосна. Конечно, первая группа пород может побиваться утренниками лишь в ненормальных для ее жизни условиях, т. е. на открытых, не защищенных пологом леса местах; здесь благодаря отсутствию защитного покрова в сильной степени может происходить излучение, которое и может повести к сильным понижениям температуры. Если в это время побеги названных пород тронулись в рост, то они легко становятся жертвами таких заморозков. В естественных условиях такие породы обыкновенно не появляются на открытых местах; обычно они живут под пологом материнского насаждения. На открытых же местах они обычно появляются без риска быть побитыми заморозками лишь после того, как такие места будут заняты пионерами древесной растительности, породами другой группы, которые не боятся весенних утренников. Когда такие открытые места после пожара или вырубки леса покроются налетом березы, белой ольхи, сосны и т. п., и когда эти пионеры более или менее сомкнутся – этим самым они как раз создадут защитный полог, который будет препятствовать сильному излучению, – тогда только породы первой группы в состоянии безнаказанно селиться под ними. Вот потому-то вероятно, что породы, боящиеся заморозков, обычно живут в условиях, защищенных от излучения, – у них и не вырабатывалось в борьбе за существование каких-либо, особенностей, которые предохранили бы их от этого зла; в их обычной нормальной жизни им просто не приходится испытывать эту особенность внешних влияний.
К ранним, или осенним, заморозкам обычно чувствительны только породы, перенесенные из более теплого климата в более холодный, привыкшие к большему теплу и к большей длине вегетационного периода, например при культуре у нас во многих местах южной части страны робинии, абрикоса и т. д. Породы же, боящиеся весенних заморозков, при переселении их в более теплый климат, еще более страдают от утренников, чем туземцы, так как, привыкнув к меньшему количеству тепла, они раньше распускаются весною, когда период заморозков еще не миновал; по наблюдениям было установлено, что наша сибирская ель больше страдает в Западной Европе от утренников, чем европейская.
Кроме низких температур древесные породы могут страдать также от высоких температур, и выражаются такие явления в виде так называемого ожога коры и опала шейки. От ожога коры могут страдать только те породы, которые имеют гладкую кору, – бук, граб, пихта, но не те, которые снабжены толстой коркой, как дуб, сосна, лиственница. Конечно, с большей чувствительностью породы могут страдать в том случае, если они, выросши в насаждении, т. е. в тени, сразу выставляются на простор. Деревья опушечные или свободно стоящие не страдают от ожога, потому что их стволы покрыты живыми ветвями почти донизу. Обращая внимание на состав пород той или иной группы, нельзя не видеть связи между возможностью ожога и теневыносливостью; породы, могущие подвергаться ожогу, с гладкой корой, суть породы теневыносливые: они гуще держатся в своих сообществах, насаждения их медленнее изреживаются; наоборот, породы, не боящиеся ожога коры даже при внезапном выставлении на простор, корковые породы, суть теневыносливые растения, которые не образуют таких плотных сообществ и насаждения которых рано и быстро изреживаются. От опала шейки страдают некоторые породы, как, например, дуб, лишь в самом молодом возрасте при росте на открытых местах и опять-таки при резкой перемене условий, при перенесении из густых питомников или тени на солнечный простор. К этой же категории явлений надо отнести и ожог хвои у ели, выставленной из-под полога материнского насаждения сразу на простор; такой угнетенный подрост имеет хвою, построенную по тенелюбивому типу, т. е. с преобладанием ткани, аналогичной губчатой. При резком переходе на свет нет времени у породы постепенным сбрасыванием старых органов создавать новые, в иных условиях освещения; в старых, не приспособленных к новым условиям, происходит разрушение хлорофилла, они погибают, а с ними гибнет и все растение.
Зимние морозы при внезапном своем наступлении причиняют трещины в коре деревьев; эти трещины обычно заплывают, но иногда наплывы вновь скрываются и опять заплывают. Значение для жизни растения такие трещины могут иметь только разве на границах распространения пород, где они, благодаря медленности заплывания, могут послужить удобным местом для поселения каких-либо паразитов. На влажных, особенно на мокрых почвах, в области черноольховых трясин например, замечается еще одно явление, связанное с низкими температурами. Это так называемое выжимание растений морозом. Происходит оно оттого, что вода, превращаясь в лед, как известно, увеличивает свой объем, отчего происходит поднятие и части рыхлой почвы вместе с живущими в ней молодыми растениями: это обстоятельство может причинять разрыв корней.
Каждая порода, занимая определенную область, встречает в пределах ее различные климатические условия. Схематически можно себе представить, что некая центральная часть соответствует оптимальным условиям, какие предъявляет порода к теплу, а две периферические части – одна по границе с более холодной областью, другая – с более теплой. В тепловом оптимуме своей области порода находит лучшие условия для своего развития – в отношении роста, плодоношения, качеств древесины, формы ствола, стойкости против всякого рода повреждений, способов вегетативного размножения, долговечности и т. д., здесь она легче всего возобновляется, является более устойчивой в борьбе со своими соперниками. Если рост какой-либо породы и становится более энергичным по мере приближения к более теплому концу своей области распространения, то такая быстрота роста не отличается продолжительностью, и если поэтому мы примем во внимание большой век дерева, то наибольшей производительностью в конечном итоге будут отличаться насаждения какой-либо породы не в более теплом, а именно в оптимальном районе своей области. То же самое справедливо и по отношению к плодоношению, которое увеличивается с передвижением к более теплому концу, – раньше начинается и чаще повторяется. Но если опять-таки мы примем во внимание время и то, что в оптимальных условиях долговечность пород больше, то в конечном итоге больше становится и тот период времени, в течение которого происходит плодоношение. В отделе, посвященном теневыносливости древесных пород, мы уже видели, что светолюбие породы не есть величина постоянная: в более теплом климате порода, как учит лесоводство, становится менее светолюбивой и, наоборот, в более холодном – более светолюбивой. Есть много и других указаний относительно различного поведения пород в разных климатических условиях: например, что породы в тепловом оптимуме своем будто бы менее разборчивы в отношении почвенных условий. произрастая же в более теплых частях своей области, требуют более влажных почв и т. п. Нужно, однако, сказать, что подобные наблюдения очень мало проанализированы, и если некоторые факты и не подлежат сомнению, то весьма часто сомнительны предлагаемые объяснения. Мы не будем поэтому сейчас останавливаться более на этом предмете. Всем, до сих пор сказанным, мы имели в виду только показать важность в жизни растений отношения их к теплу и связь, еще не вполне выясненную, но несомненную, этого фактора с другими биологическими особенностями древесных пород. Любая древесная порода представляет собой некое единство не только в морфологическом и в систематическом отношении, но и в биологическом. Та сумма свойств, которой наделена та и другая порода, не есть что-либо случайное, какая-то смесь биологических особенностей, а координированное, или согласованное, взаимное сцепление отдельных биологических особенностей, вырабатывавшихся веками при определенной внешней физико-географической обстановке и при неотъемлемом участии социальных моментов, борьбы за существование и естественного отбора.
Как внешние и внутренние органы любого организма находятся в соответствии друг с другом, так и биологические свойства представляют собой определенные гармонические аккорды. С такой точки зрения наши породы еще мало изучены, и самая классификация их совсем не разработана. Но бесспорно, что с дальнейшим развитием науки мы в состоянии будем не только лучше различать те или иные биологические типы питания, роста и размножения, но и более сложные биологические типы, образующиеся из определенного гармоничного сочетания тех или иных биологических свойств. Теперь мы знаем несколько типов лесообразователей: пионеров леса, подлесочных пород, основных лесных пород и т. п. Но со временем мы будем знать их лучше и больше: со временем, мы станем понимать, почему одни породы могут являться господствующими в насаждении, другие – всегда только входят в состав как подчиненные элементы, никогда не приобретая господства. Сейчас подмечена различная социальность роста разных древесных пород, и кое-что в этой области уж ясно, но большинство сюда относящихся вопросов еще ждет своей очереди.
БЫСТРОТА РОСТА ДРЕВЕСНЫХ ПОРОД
Биологический смысл роста в высоту – вынести вершину для беспрепятственного пользования светом. Быстрота роста имеет большое значение при борьбе пород друг с другом – как разных видов, так и в пределах одного вида. Экземпляры, наделенные большей индивидуальной силой роста, скоро приобретают господство над другими, которые при этом и переходят в разряд заглушённых и угнетенных. Быстрота роста имеет затем значение в борьбе древесных растений с живым покровом.
Энергия и продолжительность роста обусловливаются, во-первых, внутренними причинами – каждому виду свойственен свой цикл развития; во-вторых, внешними причинами – почвой и климатом; в-третьих, условиями той общественной среды, в которой растения живут, – на просторе деревья растут иначе, чем в сообществах, а в последних также различно, в зависимости от густоты и состава; в-четвертых, от происхождения, т. е.. от того, будет ли данный экземпляр семенного или порослевого происхождения (см. следующую главу).
Обращаясь к внутренним условиям, следует прежде всего отметить, что есть породы быстрорастущие и медленнорастущие. Лесоводы их располагают в ряд по быстроте роста, как, например, показывает следующая шкала, во главе которой поставлены породы с большей энергией роста: лиственница, осина, черная ольха, береза, ильмовые, сосна, клен, ясень, дуб, липа, ель, пихта. Быстрорастущие пароды – это те, которые энергично растут в молодости. Таково обычное определение. Попробуем дать более точное представление об этом биологическом свойстве пород.
Как известно, рост любого организма и любой части его подчинен так называемому закону большого периода роста. Такую кульминацию прироста в высоту, естественно, переживают и древесные породы. И вот в тех случаях, когда закон большого периода роста осуществляется в молодости, в возрасте 10 – 30 лет, породы называют быстрорастущими. Если же кульминация прироста в высоту наступает позже, породы называют медленнорастущими.
Многочисленные таксационные исследования хода роста насаждений дали многочисленные доказательства и наличности такого закона, и зависимости его от внутренних свойств пород и от внешних условий местопроизрастания.
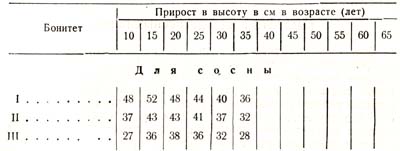
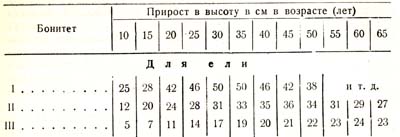
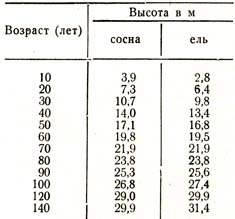
Так, оказывается, что прирост в высоту в зависимости or условий роста, достигает своей кульминации у сосны в возрасте 15 – 30 лет, у ели 20 – 50, у бука 25 – 45, у пихты 30 – 70 лет и т. д. Приведу более подробные данные для сосны и ели (табл. 23):
Таблица 23