То определение леса, которое мною дано выше, однако, недостаточно; недостаточно, не только указание на массовый момент, недостаточно даже указание на момент взаимодействия элементов в лесу; ведь и куча кирпича тоже представляет собой некую совокупность вещей, в которой тоже обнаруживается взаимное влияние кирпичей друг на друга. Для леса, как для совокупности живых тел, характерна еще другая черта, именно: устойчивость соединения древесных растений друг с другом. Каждый легко может наблюдать и на каждом шагу убеждаться, что лес обладает способностью восстановлять самого себя, если по той или другой причине было нарушено его равновесие [2 Признание в лесу равновесия, т. е. гармонии, как формы его существования взамен движения, т. е. развития леса, является идеализацией природы и механистическим представлением сущности леса. В дальнейшем в тексте «Учения о лесе» выше отмеченные утверждения встречаются часто, И они должны рассматриваться с позиций сделанных нами замечаний. Ред.] или пострадала часть его организма. Эта устойчивость происходит от способности его к самовозобновлению, тогда как куча кирпича раз разрушенная, уже неспособна сама восстановить свою целость.
Таким образом, мы будем называть лесом только такое сочетание древесных растений, которое характеризуется, во-первых, устойчивостью, во-вторых, взаимодействием друг на друга и, в-третьих, влиянием на занятую среду.
Теперь мне хотелось бы отметить те основные условия, которые делают возможной наличность общественной жизни древесных растений. Мы знаем, что последняя характеризуется прежде всего расчленением деревьев пo классам господства; некоторые стараются объяснить это явление рядом случайностей, коренящихся то в почве, то в таких явлениях, как нападение насекомых, растительных паразитов и т. д. Между тем это явление, во-первых, общее и, во-вторых, гораздо более глубокое – коренится оно в так называемой индивидуальной изменчивости. Семена ели одинакового происхождения, одинаковой величины и одинакового веса (до четвертого десятичном знака в миллиграммах), посеянные в идеально, одинаковую среду прокаленного и выщелочного соляною кислотою песка, дали всходы (рис. 13), которые по истечении двух месяцев были мною отмыты и высушены при 100°. Путем взвешивания их удалось определить, что вес сухого вещества таких растеньиц колебался в пределах 40%; так велика была между ними разница, несмотря на одинаковый вес семян и на одинаковую среду. Эр» обстоятельство коренится глубоко в недрах самого организма и составляет то явление, которое Дарвин назвал индивидуальной изменчивостью. Не будь этой коренной черты у организма, не могло бы быть и общественной их жизни. Так как, если бы все деревца росли с одинаковой силой роста, то при наступлении сомкнутости они вынуждены были бы прекратить свою жизнь. Неотъемлема потребность растений увеличивать путем роста свою деятельную поверхность, неотъемлема способность их к завоеванию пространства. Растения живут своей поверхностью, и вопрос об увеличении ее есть для них вопрос жизни; при чудовищной же плодовитости их, при оседлости наших растений и неизменности пространства единственная возможность осуществить свою потребность в росте заключается только в обладании индивидуальной изменчивостью, только благодаря ей возможна борьба между ними и как следствие – естественный или социальный отбор [1 Утверждения о чудовищной плодовитости растений при ограниченности жизненного пространства перекликаются с реакционными утверждениями Мальтуса о росте рождаемости людей в геометрической прогрессии при росте жизненных благ только в арифметической прогрессии. Академик Т Д. Лысенко показал, что в природе нет перенаселения и внутривидовой борьбы. Это необходимо иметь в виду при чтении всего курса «Учения о лесе» Морозова. Ред.]
ПОДРОСТ, ФОРМЫ и СОСТОЯНИЕ ЕГО ПОД ПОЛОГОМ ЛЕСА. ОПРЕДЕЛЕНИЕ ЛЕСА
В любом лесу, достигшем возраста возмужалости, обычно можно найти под пологом леса так называемый самосев и подрост. Они могут принадлежать к породе или породам, составляющим данный участок леса, тогда они – продукт материнского леса; но самосев и подрост могут состоять и из пород, не входящих в состав данного леса. Наличие их имеет важное значение, так как указывает на возмужалость и на возобновительную способность данного участка леса. Наоборот, его отсутствие или иной состав, чем свойственный данному участку леса, будет указывать, что существует какая-то причина, мешающая возобновлению пород, из которых он сам сложен. Подрост – это молодое поколение леса, идущее на смену старому. Количеству и качеству его поэтому лесовод не может не придавать большого значения.
Обратимся теперь к рассмотрению тех внешних форм, которыми обычно бывает представлен подрост под пологом леса. В очень молодом возрасте самосев не выделяется резко заметными особенностями, но когда он становится старше, в возрасте 10 – 30 и выше лет, смотря по породе, по характеру леса, по условиям местопроизрастания, он начинает приобретать некоторые характерные особенности в своем внешнем облике. Общей чертой, общим свойством подроста является его угнетенность. Во-первых, он всегда ниже ростом, чем одновозрастные с ним экземпляры, произрастающие где-нибудь на открытом месте по соседству. Так, например, еловый подрост под ельниками, имея высоту 1 – 1 1/2 м, насчитывает часто 60 – 80 лет, тогда как подобные экземпляры вне полога, но при совместном росте могут иметь 10 – 15 и даже более метров. Сосновый подрост, показанный на рис. 24, выдержал 24-летнее угнетение. Угнетенность роста в высоту можно хорошо видеть, наблюдая величину годичных побегов, которые тем меньше, чем больше угнетение и чем дольше живет такой подрост в этих условиях.
Рост в высоту может, однако, измениться к лучшему, как показывают рис. 25 и 26, стоит только изменить условия, например верхний полог.
Дальнейшее наблюдение «ад подростом покажет нам вторую особенность его, именно: всегда меньшее число, притом слабее развитых почек, чем у экземпляров того же возраста на свободе. В-третьих, мы здесь наблюдаем иную форму кроны, которая всегда характеризуется меньшими размерами и в особенности меньшею глубиною. У одних пород, как, например, у сосны, кроны подроста отличаются слабым, бедным и бледным охвоением, имеют чахлый вид; у других, у теневыносливых пород, как, например, у ели, пихты или бука, кроны развиваются в горизонтальном направлении, придавая при малой глубине своей зонтикообразный вид подросту и увеличивая, таким образом, поверхность прикосновения с тем небольшим количеством света, который достигает до них сквозь верхний полог леса.
Если мы, далее, срежем такие экземпляры подроста у самого основания и попробуем сосчитать годичные слои в целях определения их возраста, то будем прежде всего поражены тем, что нам вряд ли удастся это сделать невооруженным глазом. Только вооружившись хорошей лупой, и то не всегда, можно верно определить возраст подроста, который своим внешним видом, своей небольшой высотой, в пояс человека, например, невольно вводит неопытного наблюдателя в полное заблуждение. Обычно такому подросту из ели, например, неопытные люди дают 10, самое большее 15 лет, тогда как на самом деле он может иметь 40 – 60 – 80 и более лет. На прилагаемых фотографиях изображен еловый подрост из Лисинской дачи 35 лет, не достигающий и высоты человека; здесь же сфотографирован его поперечный разрез, в котором годичные слои так узки, что могли быть сосчитаны только с помощью хорошей лупы; для сравнения здесь же приложен фотографический снимок (рис. 28) с ели II класса по Крафту примерно вдвое большего возраста, т. е. 60 лет, диаметр которой, как видим, превосходит диаметр угнетенного подроста в 13 раз, а площадь сечения этого торцового разреза больше площади аналогичного сечения угнетенного подроста в 170 раз. Рис. 29 представляет собою торцовый разрез 40-летней ели из подроста (Охтенская дача Ленинградского окр.), где первые 33 года существования протекли в условиях сильного угнетения, благодаря чему годичные слои очень тонки; 8 лет тому назад было произведено изреживание, которое сейчас же сказалось на расширении годичных слоев. На рис. 30 показан поперечный разрез через стволик сосны из подроста 24-летнего возраста, где тонкость годичных слоев и последующее их расширение также хорошо заметны. Еще лучше это видно на рис. 25, представляющем так называемый таксационный анализ ствола и дающем представление о ходе роста в высоту и в толщину этого экземпляра. Рис. 31 показывает ель, оправившуюся после 160-летнего угнетения материнского полога.
Указанные обстоятельства, т. е. задержанный рост в высоту и в особенности в толщину, влекут за собою и незначительное развитие объема стволиков подроста по сравнению с экземплярами того же возраста, выросшими на той же почве, но вне влияния полога леса.
Ко всему сказанному необходимо прибавить, что корневая система подроста, в соответствии, конечно, с развитием надземных частей, отличается тоже более слабым развитием, меш шей деятельною поверхностью и меньшей глубиной проник но вения в почву.
Хвоя и листья, заложенные в тени, несут также в своем анатомическом строении печать тех условий, в которых они

Рис. 25. Схема поперечного разреза подроста
родились. Такая хвоя, будучи приспособлена к работе при малом количестве света, при выставлении на простор желтеет, отпадает, и деревцо или подрост в таких случаях, по народному выражению, «пугается»; необходимо такое выставление на простор произвести постепенно, пока не сменится у дерева хвоя.
В других случаях подрост гибнет при внезапном выставлении на простор от другой причины – он засыхает от недостатка влаги. Происходит это не потому, что на открытом месте по удалении леса в почве будет меньше влаги, наоборот, влаги будет больше, но ею слабо развитая корневая система воспользоваться не может; подрост под пологом леса окутан более влажной атмосферой, в гораздо меньшей степени подвержен ветру, пользуется меньшим количеством света и тепла и в силу всего сказанного испаряет весьма мало; будучи же выставлен на простор, он принужден испытать в резкой форме перемену условий; и большая инсоляция, и больший доступ ветра, и более сухой воздух могут чрезвычайно сильно повысить его физиологическое испарение, между тем его корневая система, приспособленная к иным условиям, может оказаться не в состоянии удовлетворить этому большему запросу на влагу. Иногда это кончается очень трагично – быстрым засыханием подроста; иногда такая агония длится много лет, и в результате растение все равно погибает или только от этой причины, или потому, что такой ослабленный организм добивается вторичными врагами – насекомыми, например. Но в некоторых случаях после долгой борьбы за жизнь такой подрост начинает оправляться. Зависит это, конечно, от многих причин, но главным образом, с одной стороны, от степени угнетенности его, с другой – от степени внезапности выставления на простор;

Рис. 28. Торцовый разрез ели 63 лет, выросшей в насаждении в качестве дерева II класса по господству

Рис. 29. Торцовый разрез оправившегося елового подроста. Возраст 35 лет
затем, конечно, от возраста породы, условий местопроизрастания, погоды и т. п.
В тех случаях, когда укоренение подроста весьма поверхностно, в особенности же когда часть корней даже живет в слое подстилки, обычно прикрывающей в лесу почву, засыхание подроста при внезапном выставлении на простор может происходить и от большей сухости этих поверхностных горизонтов, которые в отличие от более глубоких слоев почвы и грунта на открытых местах могут быть суше, чем в лесу. Большая сухость этих верхних горизонтов может, наконец, происходить от задернения травяным покровом – сильным потреби-

Рис. 30. Торцовый разрез соснового подроста, 19 лет находившегося в угнетении и оправившегося после изреживания верхнего полога. По данным проф. Соболева (натур, величина)

Рис. 31. Ель, оправившаяся после 160-летнего угнетения, по изреживании верхнего яруса 30 лет назад
телем влаги, могущим поэтому явиться соперником в этом отношении древесной растительности.
Во многих случаях у чувствительных к заморозкам пород, например, у ели, пихты и бука, подрост при внезапном выставлении может погибнуть прежде всего от заморозков, так как на открытых местах такие деревца ничем не защищены от излучения, а весною вегетационный период здесь начинается раньше, чем под пологом леса, который защищает подрост от излучения.
Познакомившись с характерными особенностями внешнего вида подроста, его угнетенностью, с своеобразной формой кроны, малыми размерами и пр. и пр., естественно поставить вопрос, отчего это происходит? Отчего древесные растения одного и того же возраста, произрастающие вне полога леса, но тоже в борьбе за существование с себе подобными, так сильно отличаются по своим размерам в высоту, в толщину и по объему от растущих под пологом леса? Очевидно, потому, что подрост под пологом всякого леса, – все равно, материнского или из других пород, – испытывает двойную конкуренцию, двойную борьбу за свое существование. В группах подроста происходит та же дифференциация, о которой шла речь в предыдущей главе и которая свойственна всякому лесу без исключения, но кроме того здесь происходит борьба такого подроста с материнским организмом или тем лесом, который приютил его под своим пологом. Подрост под пологом леса пользуется меньшим количеством света, так как падающий рассеянный свет прежде всего используется пологом материнского насаждения или (верхнего яруса, а затем уже тот, который отработал в листовых органах верхнего полога, в смеси с тем, еще не работавшим, рассеянным светом, который проникает сквозь отверстия верхнего полога леса, достается на долю потомства. Подросту затем приходится распространять свои корни в почве, занятой уже корнями дерев верхнего яруса, и приспособляться к условиям, созданным в этой среде материнским или защитным насаждением.
Не испытывая конкуренции с травянистою растительностью, пышно не развивающеюся под пологом леса, подросту приходится соперничать с корнями верхнего яруса в отношении влаги.
Приведенные данные и соответствующие фотографии показали, что изреживание верхнего полога улучшает рост подроста, ослабляет его угнетенность. Отчего это происходит? Очевидно, от ослабления конкуренции, от ослабления жестокой борьбы за существование между подростом и верхним пологом. Если это ясно, то всплывает другой вопрос: в каком же направлении ослабляется конкуренция? Вообще говоря, на этот вопрос ответ может быть один: во всех направлениях, так как, на самом деле, изреживая верхний полог, мы 1) увеличиваем доступ света к подросту, доступ тепла и осадков, которые теперь меньше будут задерживаться кронами дерев верхнего яруса, 2) ослабляем конкуренцию корней в почвенной среде, 3) увеличиваем благодаря этому и большему доступу осадков влажность почвы и грунта, 4) изменяем условия разложения подстилки и гумуса и т. д. В отдельных же случаях можно иногда указать, какой из перечисленных факторов является решающим: у светолюбивых пород в районах с большой влажностью воздуха, с большой облачностью, с большим количеством пасмурных дней, на влажных почвах улучшение условий жизни подроста под влиянием изреживания верхнего полога может быть приписано увеличению доступа света, как, например, для сосны Ленинградской области, Брянского массива Орловский окр. В противоположных условиях, в сухом климате нашего юго-востока, притом в сухих местоположениях, причиной улучшенного роста подроста при изреживания насаждения может быть большее количество влаги в почве благодаря ослаблению конкуренции с материнскими корнями. В лесоводстве были произведены соответствующие опыты, доказавшие справедливость сказанного; мы их изложим в свое время подробно, теперь же упомянем только о том, что, не изреживая верхнего полога, а лишь окапывая группы соснового подроста канавками (в которых обрубались все материнские корни, после чего канавки вновь засыпались), изолируя, таким образом, корни подроста от корней старых деревьев, удавалось, во-первых, увеличить запасы влаги в почве, во-вторых, улучшить рост подроста по сравнению с теми группами его, которые не подверглись указанной операции.
Итак, подрост есть продукт всей своеобразной обстановки под пологом леса, то более, то менее чувствительный реактив на степень энергии происходящей здесь борьбы за существование. Подрост представляет собою продукт наибольшего социального гнета по сравнению с тем угнетением, которое испытывают древесные растения одного и того же яруса в лесу.
Полагаю, что все сказанное о подросте присоединяет к прежним двум группам фактов достаточно солидные доказательства того, что лес не есть простая совокупность древесных растений, а представляет собою сообщество [1 Обозначение леса как совокупности взаимно связанных в своей жизни растений словом «сообщество», применявшееся Морозовым, при критике его идеологических ошибок в 30-х годах было отвергнуто в связи с тем, что оно сближает лес с человеческим обществом. Взамен русского слова «сообщество» были предложены различные греческие и латинские названия, в частности слово «фитоценоз», представляющее комбинацию двух греческих слов: «фитон», что означает растение, и «ценоз» – искаженное слово «коииоз», означающее в переводе общество. Все вместе, следовательно, означает общество растений. Нельзя признать целесообразной замену русского обозначения растительной группировки «сообщество», которое хотя и близко к слову «общество», но подразумевает отличия от него, комбинацией греческих слов «фитоценоз», которое уже стирает разницу между растительной группировкой и человеческим обществом и приводит к так называемому социальному дарвинизму. Ред.]
или такое соединение древесных растений, в котором они проявляют взаимное влияние друг на друга, порождая тем целый ряд новых явлений, которые не свойственны одиноко растущим растениям.
ИЗМЕНЕНИЕ ВНЕШНЕЙ СРЕДЫ ПОД ПОЛОГОМ ЛЕСА. ПОДСТИЛКА
Указанными признаками не исчерпывается, однако, сущность того соединения древесных растений, которая именуется лесом. Всем хорошо известно, что в лесу, или, еще точнее, под пологом леса, другой климат, другая почва и иной почвенный покров. Полог леса (рис. 32), подобно продырявленному зонтику, с которым иногда его сравнивают, пропускает сквозь себя не все количество осадков, выпадающее в данном пункте земли; часть осадков, смачивая хвою, листья, ветви и т. д.,
остается на кронах, откуда затем испаряется обратно в атмосферу, другая часть сквозь отверстия в пологе непосредственно проникает допочвы и, наконец, третья стекает по стволам к подошве дерева. Точные и многочисленные измерения, о которых у нас речь будет в особой главе, показали, что полог леса задерживает от 15 до 80% на своих кронах, в зависимости от породы, густоты Древостоя, возраста леса и других моментов. Так, сосновые леса задерживают только около 20%, еловые около 40%, а пихтовые около 60% от количества осадков, выпадающих на открытом месте. Величина стока по стволам у одних пород совсем незначительна – менее 1%, у других, как у бука, наоборот, довольно значительная, достигает 5%.
Что говорят нам только что приведенные данные, детальное рассмотрение которых нам еще предстоит? Они говорят нам о том, что если у нас на единице площади земли выпадает в течение года, положим, 500 мм осадков, то стоит нам развести лес, как количество осадков, которое будет достигать нашей почвы, упадет на 20 – 40 – 60%, в зависимости от породы, которую мы выберем при образовании леса. Таким образом, 100, 200, 300 мм осадков будут изъяты из общего баланса влаги данного места. Это значит далее, что и сами деревья, которые будут образовывать лес, и их потомство, которое со временем возникнет у них в виде подроста под пологом, будут пользоваться меньшим количеством осадков, чем свойственно данному географическому положению. Пока деревца, которые мы насадили на нашей площади для образования леса, еще не сомкнулись своими кронами, пока они представляют собою простую совокупность хота и большого множества деревьев, но в одиночном стоянии, до тех пор лесная почва будет пользоваться почти полным количеством осадков, выпадающих в данном месте. Но как только деревца сомкнутся, для них и их потомства начинается новая жизнь. Сквозь полог будет уже меньше проникать света и тепла, и созданный лес будет препятствовать проникновению внутрь его ветра. Благодаря этому под пологом леса складывается иной тепловой и световой климат, иная относительная влажность воздуха и т. п. Подрост под пологом будет помещен в иные условия влажности и испарения и т. д. Это видоизменяющее элементы атмосферы влияние леса не ограничивается сказанным, а проникает и глубже в другую среду, именно почвенную, где также умеряет тепловые амплитуды и в общем среднем делает лесную почву более холодной.
Но для того, чтобы оценить преобразующее влияние леса на почву, необходимо обратить внимание на одно весьма характерное явление, присущее лесу. В лесу всегда имеется, кроме живого покрова из теневыносливых трав, мхов и т. п., еще так называемый мертвый покров, или подстилка, которая составляется из опавших листьев, мелких веток, погибших семян и тому подобных растительных остатков. Из общего курса ботаники мы знаем, что листовые органы служат для ассимиляции углерода, для дыхания и для испарения; в лесу же к ним, как заметил лесовод Бюсген, присоединяется еще новое отправление: лист, сбрасываясь, образует подстилку, которая, разлагаясь в определенных условиях лесного климата, производит глубокие изменения в нижележащей почве. Почвоведение показывает нам, какую существенную роль продукты разложения этой подстилки играют в том подзолообразовательном процессе, который свойственен всем лесам наших климатических областей. Если лес является крупным почвообразователем, то главным образом благодаря этой подстилке и тем климатическим условиям, которые создаются левом под его пологом и которые определяют характер перегнивания этого «мертвого покрова.
Эта подстилка является источником гумуса лесной почвы, влияет в качестве мертвого покрова на физические свойства нижележащей почвы, образует ближайшую среду, в которой приходится прорастать семенам древесных растений, а при некоторой толщине подстилки и прозябать в ней в сходам самосева и даже подросту.
Сложный механизм влияния подстилки на почву, на живой покров и на возобновление леса будет также рассмотрен в своем месте; сейчас же мы ограничимся только сказанным и в заключение подчеркнем еще раз, во-первых, громадное и весьма разнообразное влияние ее на указанные выше обстоятельства и, во-вторых, что подстилка представляет собою новую биологическую среду, созданную притом лесом.
Помимо подстилки и тех атмосферных условий, которые создает лес под своим пологом, он влияет на почву и непосредственно своими корнями, которые изменяют структуру почвенной среды и, высасывая в громадных количествах влагу для нужд физиологического испарения, иссушают почву и грунт под лесом. Многочисленные исследования влажности почвенной среды под лесом, произведенные у нас в стране и в Западной Европе, – все без единого исключения доказывают один основной факт, что почва под пологом леса только в самых поверхностных горизонтах, несколько они не пронизаны корнями, влажнее под лесом, чем вне его; более же глубокие горизонты, в которых помещается корневая система верхнего яруса, подлеска и т. п., суше соответствующих горизонтов земли, не покрытых лесом. И этот весьма важный и сложный вопрос будет в свое время весьма подробно разобран с критическим освещением всех относящихся сюда источников.
Собирая все сказанное в этой главе, мы в состоянии сделать тот логический вывод, что обстановка жизни под пологом леса иная, чем вне его, что местный климат и почва изменяются лесом и, как мы увидим впоследствии, изменяются именно в зависимости от состава леса, его возраста, густоты древостоя и т. п. Но ведь в эту новую, до известной степени преобразованную, климатическую и почвенную среду погружены те самые растения, которые ее создают; в еще большей мере испытывает на себе влияние этой новой обстановки созданный лесом или просто поселившийся под его пологом подрост.
В этой новой, измененной жизненной обстановке – ключ, как мы увидим впоследствии, к уразумению [1 Значение структуры почвы как решающего фактора плодородия глубоко раскрыл академик В. Р. Вильяме, который показал также возможность и необходимость изменения структуры почвы путем смены однолетних растений многолетними и наоборот – многолетних однолетними в форме так называемой травопольной системы земледелия Ред.] особенностей формы, роста и других сторон жизни дерев, образующих лес, в отличие от свободно растущих. В этой же новой обстановке – ключ к уразумению судеб подроста, его появления и исчезновения, особенности его; роста и т. д., и т. д. В этой новой обстановке лес, как увидим впоследствии, приобретает новое средство борьбы за существование; светолюбивые элементы лесной флоры, не будучи в состоянии мириться с тем малым количеством света, которое бывает под пологом леса, не могут проникнуть внутрь участков из теневыносливых пород и в своей вечной жажде к завоеванию новых пространств принуждены довольствоваться открытыми, безлесными местами, где они легче одолевают сопротивление травяного покрова. Известных качеств подстилка и производный из нее гумус могут привлекать одну породу и не отвечать потребностям другой. Одним словом, в этой новой обстановке, созданной и постоянно создаваемой и поддерживаемой лесом, как увидим впоследствии, мы найдем ключ к уразумению лесных метаморфоз и динамики леса.
Прежнее определение леса расширяется; к прежним существенным признакам может быть прибавлен новый, столь же существенный, как и прежние. Лесом мы будем называть такую совокупность древесных растений, в которой обнаруживается не только взаимное влияние их друг на друга, но и на занятую ими почву и атмосферу.
ЗАЩИТНОЕ ВЛИЯНИЕ ПОЛОГА ЛЕСА
Древесные растения в лесу, как мы видели, находятся во взаимодействии, причем с одной стороной этой взаимной связи мы познакомились в виде борьбы за существование древесных растений друг с другом. Это самая главная и лучше всего изученная сторона указанного явления, но нельзя не видеть, что взаимодействие между деревцами имеет еще другую сторону противоположного характера, которая известна в лесоводстве под именем защитной способности пород и древесного полога. То самое соединение древесных растений, которое порождает борьбу за существование и создает, как мы видели в прошлой главе, своеобразную обстановку, заключает в себе также и защитные элементы. Уже было обращено, внимание на то, что под пологом леса не бывает пышно развитого травяного покрова, теневыносливые представители лесной флоры рассеяны довольно редко и ни надземными, ни подземными своими частями не могут представлять серьезных соперников древесным всходам. Прилагаемая фотография показывает ту мощную травяную, чисто злаковую, растительность, которая часто развивается на открытых пространствах возле леса, с которой приходится соперничать появляющимся среди нее древесным всходам. На двух следующих фотографиях показаны отдельно дернина вейника, с одной стороны, и густое сплетение корней этой дернины – с другой. Такой мощный травяной покров и сильно сушит почву, и может вредить усилением заморозков над своей зеленой поверхностью, может заглушить древесные всходы тенью, может затруднять проникновение
их корневой системы к почве благодаря толстому слою войлока из отмирающих его надземных частей. Следующие фотографии
показывают, как фактически различие в характере травяного покрова отражается на росте деревьев. Мы видим, что те же 5-летние дубки, выросшие среди злаковой растительности, в несколько раз ниже ростом и тоньше, чем одновозрастные с ними дубки, росшие среди гораздо более редкого травостоя из сныти (Aegopodium podagraria,).
Сказанное (показывает нам, что в пологе леса действительно заключается известный защитный элемент, который состоит в том, что он благодаря своей тени устраняет травяную светолюбивую растительность, устраняет этих конкурентов как для самого, себя, так, в еще большей мере, для своего потомства.
Мы видели также из прежнего изложения, что полог леса защищает от излишней инсоляции, а также от чрезмерного излучения те древесные растения, которые поселяются под ним; благодаря этому чувствительные к заморозкам деревца не страдают под защитою леса от утренников. Когда сплошь срубается часть елового леса, то обыкновенно на таких открытых местах, быстро заселяющихся травами, ель сразу не поселяется, хотя бы возле была плодоносящая стена елового леса. И не то, чтобы всходы, ели не появлялись, наоборот, внимательное наблюдение таких открытых мест всегда может доказать противное. Среди травы мы легко найдем всходы, а иногда и подрост ели, погибающий или погибший от заморозков. Через некоторое число лет, однако, такая площадь вновь завоевывается елью, в чем можно убедиться на любой экскурсии в еловый лес. Как же это происходит? Если мы возьмем такие вырубки разного: возраста, то в состоянии будем заметить следующую последовательную картину. Лесосеки покрываются березовым или осиновым налетом. Указанные породы обладают чрезвычайно легкими семенами, их всходы – быстрым ростом, относительной нечувствительностью к заморозкам; благодаря такому сочетанию своих биологических или лесоводствеиных свойств эти породы приспособлены к завоеванию открытых мест, так как быстро оправляются с травою и не боятся заморозков. Когда этот налет сомкнётся, когда такой налет создаст лесные условия и под собою тень и защиту от излучения, тогда начинает под таким защитным пологом селиться чувствительная к заморо1экам ель. С одной стороны, такой полог из березы и осины, а иногда и других пород (белой ольхи, сосны), будет отнятием света и пр. до известной степени угнетать сидящий под ним еловый подсед; с другой – не будь этого полога, не появился бы этот подрост из ели. Мы видим несомненную защитную способность лесного полога и видим в то же время, что угнетающее и защитное влияние полога – две стороны одной медали, что, они неразрывно связаны и, наконец, что в данном случае защитное влияние имеет большую биологическую или жизненную ценность, чем конкурирующее влияние полога, которое должно рассматриваться как неизбежный спутник влияния первого рода.
Что будет происходить в дальнейшей жизни таких двух лесных сообществ? Березовое насаждение в силу закона борьбы за существование будет постепенно изреживаться; среди него, как среди любого леса, будут постепенно появляться угнетенные классы; число стволов у березы, как у светолюбивой породы, будет убывать быстро, и такое защитное насаждение будет с годами пропускать больше света, тепла, осадков и меньше угнетать ель. Еловый подсед, постепенно поднимаясь выше и выше, с течением времени превратится во второй ярус, а затем, со следами угнетения, с большим уроном для себя, после жестокой двойной борьбы за существование, начнет постепенно врастать в состав верхнего яруса. Ель – порода медленно, но длительно растущая и теневыносливая; береза, наоборот – светолюбивая, менее долговечная и хотя и быстрорастущая, но гораздо раньше ели прекращающая свой рост в высоту. Вот, когда ель достигнет верхнего полога, тогда плохо придется березе; если ель, хотя и с уроном для себя, долгое время могла сносить отенение березы, то последняя, очутившись под елью, уже не в состоянии будет помириться с этим и начнет отмирать; ель в это время совсем перегонит бывших своих покровителей, и вновь восстановится еловый лес с небольшою примесью мягких лиственных пород (березы и осины). Но стоит вновь вырубить такой еловый лес, и природа снова опишет этот же круг.
До чего реальна эта защитная способность березового полога, свидетельствуют не только: факты, которые были только что сообщены, но и то обстоятельство, что лесоводственные деятели на практике использовывают эту защитную способность. Когда нужно на какой-нибудь голой площади, которую усердно посещают весенние заморозки, развести лес из чувствительной к заморозкам древесной породы, например ели, то лесничие раньше создают культуру какой-нибудь второстепенной, не боящейся заморозков породы, например березы, и когда такое насаждение начнет смыкаться, то под его полог вводят ценную породу. Лесничие, как видим, подражают природе.
В сосновых лесах в виде подлеска встречаются кустарники из семейства мотыльковых; они имеют большую биологическую ценность для соснового леса как азотособиратели. Эта особенность их использована в практике лесоводства с большим успехом; там, где при создании елового леса вводили одновременно в междурядия такие кустарники, там культура удавалась лучше и рост ели выгодно отличался от мест, где не было этих сотрудничающих элементов.
В лесоводстве известно много случаев, показывающих, что некоторые породы лучше уживаются, лучше возобновляются, лучше культивируются, когда бывают в сообществе с другими породами, а не тогда, когда они растут в чистых насаждениях.
Этим я ограничусь в характеристике защитных способностей леса, так как вопрос этот и чрезвычайно сложен и весьма мало еще разработан; в свое время мы на нем остановимся, сколько будет нужно, сейчас же моя цель исчерпана, так как я хотел только отметить наличность другой стороны, другого элемента в том взаимодействии между древесными растениями, которое приковало наше внимание с момента нашего вступления под своды леса.
НЕКОТОРЫЕ ИЗМЕНЕНИЯ АНАТОМИЧЕСКОГО СТРОЕНИЯ ДРЕВЕСИНЫ ПОД ВЛИЯНИЕМ СОЧЕТАНИЯ ДРЕВЕСНЫХ РАСТЕНИИ В СООБЩЕСТВО
Взаимное влияние дерев в лесу не ограничивается изменением внешней формы и роста их, но, распространяется и на внутреннее строение их. Удельный вес древесины, состав ее и анатомическое строение, которым обусловливается плотность ее, все это различно в пределах одной и той» же породы на одной и той же почве, в одном и том же климате, в зависимости лишь от того, выросло ли дерево на свободе или в насаждении, а в последнем случае – к какому классу господства оно относится. Давно в практике было известно, что хвойные деревья, выросшие на просторе, отличаются менее плотной древесиной, чем в сомкнутых насаждениях, что удаляемые при прореживаниях угнетенные стволы ели обнаруживают более плотную и более прочную древесину, чем классы господствующие, что выросшие под защитой материнского насаждения ели и пихты обладают древесиною выдающихся качеств столетних насаждениях пихты и ели удельный вес абсолютно сухой древесины, по исследованиям Бертога [1 По вопросу о зависимости свойств древесины от положения дерева В насаждении имеются отечественные исследования Яхонтова, Савиной и других. – Ред.], достигал максимума у дерев III класса, уменьшаясь как в сторону классов большей степени господства, так и в сторону угнетенных, как показывает это табл. 6.
Таблица 6

Тот же автор обнаружил при этом, что отношение толстостенной части годичного кольца, осенней, к тонкостенной или весенней, за последние 50 лет у ели, притом у разных классов господства, выразилось следующими величинами:
VI кл. – 0,36:1; III КЛ. – 0,55:1; IV кл. – 1:1.
В связи со степенью господства находится и величина трахеид.
Толщина стенок трахеид, будучи измеренной в микронах (в тысячных долях миллиметра), варьирует, например, у ели таким порядком: 1 – 4,4; II – 4,5; III – 4,3 и IV – 3,7.
Площадь поперечного сечения трахеид изменяется в зависимости от степени господства следующим образом, в квадратных микронах (в миллионных долях квадратного миллиметра):

Мы видим, что по мере угнетения размеры трахеид становятся меньше, что не может не иметь влияния на удельный вес древесины, в данном случае увеличивая ее плотность.
Другое исследование Р. Гартига дает нам для 100-летней ели на высоте 1,3 мм следующие данные о тангентальных диаметрах трахеид и о площадях поперечного сечения их (табл. 7):
Таблица 7

Угнетенные стволы характеризуются меньшими размерами клетки, причем эта разница достигает таких размеров, что в одной трахеиде господствующего класса могут поместиться три трахеиды наиболее угнетенного.
Содержание лигнина в единице веса больше у дерев, пользовавшихся простором, чем у угнетенных, причем колебания эти бывают больше, чем различия, обусловленные породами:
Ель на просторе: 0,93 – 1,07 – 1,02%
Ель угнетенная: 0,84 – 0,73 – 0,87%
Аналогичные данные приведем еще для другой породы – для дуба. Здесь точно так же Р. Гартиг нашел, что плотность древесины зависит, помимо всех прочих условий, и от степени угнетения стволов; так, в 100-летнем дубовом насаждении средний вес 1 плотного м3 древесины (в воздушно-сухом состоянии) у дерев разного класса колеблется следующим образом:
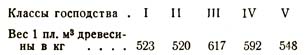
В другом примере он нашел, что в 100-летнем дубовом насаждении средний вес 1 м3 древесины (в воздушно-сухом состоянии) изменялся в зависимости от класса господства следующим образом: /
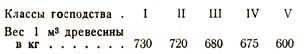
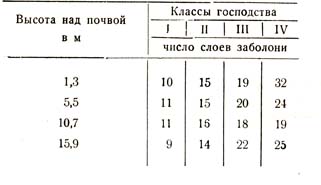
Тот же исследователь показал, что и число слоев заболони колеблется в определенном направлении у представителей различных классов господства, именно: оно увеличивается по мере угнетения стволов, как показывают следующие данные для 95-летнего дубового насаждения (табл. 8).
Таблица 8
Количество сердцевинных лучей тоже колеблется в зависимости от интересующего нас момента; так, на высоте груди в 140-летнем дубовом насаждении сердцевинные лучи занимают: у экземпляров I класса – -8,4%, у II класса – 6%, у III класса – 3,7% поверхности среза древесины; у 400-летнего дуба, стоявшего на просторе с давних пор, оказалось наибольшее количество сердцевинных лучей, именно 10,4%.
В другом примере Р. Гартиг дает еще большие разницы, а именно: у дерев на просторе означенный выше процент равнялся 11 – 12, а у дерев, выросших в сомкнутости – только 4,6.
Я могу ограничиться приведенными примерами, так как вовсе не имею в виду исчерпать имеющийся фактический материал, ни даже его систематизировать; я хочу только обратить внимание на то, что он существует, и на то, что он показывает; а показывает он, что анатомическое строение древесины зависит не только от породы и внешних условий роста (климата, почвы), но и от того, насколько сомкнуто насаждение и к какому классу господства относится данный экземпляр. Эти и другие не приведенные аналогичные данные хорошо разъясняют, что при изучении древесины, ее качеств и строения, так же точно, как и при исследовании роста, формы и плодоношения деревьев в лесу, нельзя это осуществить, не обращая внимания на классы господства, что наряду с видовыми признаками и теми особенностями, которые обусловливаются внешней средой, необходимо считаться и с теми, которые обусловливаются другой добавочной, создаваемой самим лесом – внутренней средой – средой социальной.
Как бы там ни было, права ли эта теория во всем объеме или она нуждается в дополнениях, налицо остаются бесспорные факты, давным-давно известные практикам, давно уцененные лесоводством, показывающие, что анатомическое строение древесины зависит не только от видовых особенностей и внешней обстановки, но и социальной среды, в которую древесные растения погружены, которую они создают и в которой они занимают различное общественное положение. Претворенная совокупностью древесных растений внешняя физико-географическая обстановка и является той новой средой, под влиянием которой меняются не только формы деревьев, но и их внутреннее строение.
На основании сказанного, мы можем видоизменить определение леса, какое мы раньше давали, мы можем понимать под лесом совокупность древесных растений, измененных как в своей внешней форме, так и в своем внутреннем строении под влиянием воздействия их друг на друга, на занятую почву и атмосферу.
РАЗЛИЧНЫЕ ФОРМЫ ЛЕСА
До сих пор шла речь о существенных признаках леса, притом объединяющих, т. е. свойственных всякому лесу. Теперь обратимся к другого рода признакам, я бы сказал разъединяющим, т. е. таким, по которым можно отличить один лес от другого. Лес ведь на мало-мальски большом пространстве почти никогда не бывает однородным. Неопытный наблюдатель на первых порах теряется в лесу; ему обычно в нем представляется все чересчур разнообразным, чуть не с каждым шагом он связывает перемены в лесу; нужен некоторый опыт выделять однородные в себе части, отличая их от соседних; надо знать признаки, по которым выделяются однородные в себе и отличные от соседних части леса. Такими признаками, очевидно, могут быть только такие обстоятельства, которые определяют общественную жизнь древесных растений в лесу. К ним относятся прежде всего признаки внешние, т. е. различия в условиях местопроизрастания, во-вторых, признаки внутренние, принадлежащие самому лесу. Как лицо мало знакомое с ботаникой не в состоянии дать представления о растении, им виденном, так и большинство посетителей леса, незнакомых с ним, не в состоянии охарактеризовать те лесные участки, которые они посетили; они могут сказать, что были в сосновом лесу, добавить еще к этому несколько случайных верных указаний, а затем пойдут уже совсем ненужные слова, вроде: дремучий, темный, глухой и т. п. Подобные термины так же мало могут дать для верного; представления о посещенном лесе, как и такие выражения, что «цветочек был такой маленький и синенький» и т. п. Так по каким же внутренним признакам можно отличить одно лесное сообщество от другого?
Первый вопрос, в котором надо себе отдать отчет, наблюдая какой-нибудь лесной участок, – это занята ли данная площадь земли одним или несколькими лесными сообществам»; если над почвой возвышается всего только один полог, то это будет одноярусная, или простая, форма леса; если же над почвой возвышаются два и более пологов, то перед нами сложная, или много-ярусная, форма лесного насаждения. На прилагаемых фотографиях изображены: одноярусное насаждение из бука; двухъярусное – в виде дуба с подлеском из татарского клена. Насаждения, в состав которых входят различные породы, мы называем смешанными; состав еще больше разнообразит картину леса. Надо добавить, что подлеском называется тот случай, когда нижний ярус состоит из кустарников или из древесных пород, но которые при данных почвенных условиях принимают кустарниковые размеры.
В числе признаков, о которых сейчас идет речь, я поставил на первое место так называемую форму леса, или форму (насаждения, так как, ведь на самом деле, надо же прежде всего знать, имеем ли мы дело с одним или со многими сообществами, живущими на одной и той же территории и возвышающимися друг над другом. В случае сложного насаждения каждый ярус описывается отдельно по признакам, о которых сейчас и будет итти речь.
Вторым вопросом является характеристика каждого яруса, если насаждение сложное, и одного яруса, если оно простое, в отношении состава пород, образующих наблюдаемый участок земли. Лесоводство обыкновенно не довольствуется одной только качественной характеристикой состава леса, т. е. перечнем пород в порядке их господства, или выделением господствующих элементов от подчиненных, а требует количественной характеристики, указывая верно степень участия каждой породы в составе леса. Особенностью лесоводства при этом является то обстоятельство, что степень участия характеризуется по числу деревьев той или иной породы на единицу площади, а доле участия каждой из них в общем древесном запасе или объеме древесины, образуемой ярусом, на единице площади. В результате может получиться такая формула: 8/10 дуба, 2/10 ясеня; или 6/10 ели, 2/10 Сосны, 1/10 Березы, 1/16 Осины, или еще: 7/10 Сосны, 2/10 Ели, 1/10 Березы + Осины, когда последняя составляет менее 1/10, т. е. когда порода попадается единично, или, как говорят, вкраплена. Все десятые в сумме, конечно, должны дать 1; десятых можно не писать, ограничиваясь помещением одних числителей; вместо названия пород можно помещать одни лишь начальные буквы; формула тогда становится компактнее и может (Приобрести, например, следующий вид: 6Е,2С,2Б + Ос.
Но надо было прежде всего сказать, что если в отношении формы мы различали простые насаждения от сложных, то в отношении состава лесоводство различает насаждения чистые от сообществ смешанных. Между ними, естественно, есть целый ряд переходов, так что во многих случаях граница будет носить условный характер. Чаще всего считают, что если участие какой-нибудь породы в составе господствующей части будет менее 2/10, то насаждение будет считаться чистым, если более 2/10, то смешанным.
В природе часто встречаются сложные насаждения, в которых каждый ярус или некоторые из них представляют собою смешанные сообщества.
Каждый ярус, каждое насаждение должно затем характеризоваться в отношении возраста, причем различают прежде всего насаждения одно возрастные и разновозрастные. Если разница в возрасте у древесных растений, входящих в насаждения, не превышает известной условной величины, так называемый состав класса возраста, то такое лесное сообщество будет называться одновозрастным, в противном случае – разновозрастным; обычно классы возраста делают 20-летними, иногда 10-летними. В случае одновозрастности насаждения оно относится к I, II, III и т. д. классу. Возраст обыкновенно пишут в скобах и помещают следом за названием породы и степенью ее участия в составе насаждения, например 9/10 (100 – 120 л.), 1/10С (120 – 140). Иногда пользуются естественными ступенями для характеристики возраста насаждения, причем под молодняком или чащей (разумеют насаждения до начала заметного, расчленения на классы господства; под жердняком – от начала деятельного переживания до конца этого периода; с затиханием этого, процесса, с наступлением возмужалости лес становится приспевающим, а затем спелым. Эти термины в силу своей неопределенности могут затруднять пользование ими на практике.
Следующий элемент, который должен быть охарактеризован при описании лесных сообществ, – это густота или полнота насаждения и степень ее равномерности в пределах описываемого участка. Под этим термином разумеют два понятия: степень сомкнутости крон – это одно, густота древостоя – это другое. Понятное дело, что: обыкновенно с увеличением числа дерев, или плотности древесного населения, увеличивается и степень сомкнутости полога, но насаждения могут быть вполне сомкнуты своими кронами и при меньшем числе стволов, если кроны у последних хорошо развиты. Насаждения, в которых кроны деревьев соприкасаются друг с другом, так что их проекции на почву покрывают и, стало быть, отеняют ее всю, называются полными. Для выражения степени полноты пользуются тем же принципом, как характеризуется облачность в метеорологии; обозначая 1,0 насаждения полные, определяют затем степень полноты десятичною дробью в зависимости от того, какую часть площади занимает проекция крон. Иногда довольствуются только1 тремя обозначениями: насаждения полные, средней полноты и редкие. Что же касается густоты древостоя, то ею пользуются лишь при таких точных описаниях, когда производится предварительный перечет всех деревьев на пробных площадях.
За отметкой полноты дается указание относительно средней высоты господствующих деревьев (II класса) насаждения. Такая средняя высота, определяемая на глаз или с помощью инструмента, при умелом пользовании ею может дать хорошие указания относительно условий роста, служа как бы масштабом для сравнительной характеристики условий местопроизрастания. Можно отмечать также и средний диаметр господствующих в насаждении деревьев. Лесоводы обычно вводят в описание указание на прирост, на общую древесную массу и тому подобные элементы так называемого таксационного описания, о, котором речь сейчас итти не может, так как для этого необходимы специальные познания.
Весьма важно отметить состояние насаждения, поскольку оно еще не выразилось в полноте; это – «степень его совершенства при данных условиях местопроизрастания», сюда относится указание о степени здоровья деревьев и о их фаутности, О) чем можно судить часто по наружным признакам: по грибным повреждениям, по чрезмерному развитию лишайникового покрова на стволах деревьев, по суховершинности, по цвету хвои или листвы, по ненормальностям в развитии кроны, иногда стволов и т. п.
Таких примеров я бы мог, конечно, привести великое множество, и мы с ними будем иметь еще много дела на нашем долгом пути познания природы леса. Я думаю, что если бы привел еще несколько примеров, то едва ли я увеличил бы надежность того вывода, который, полагаю, достаточно ясен и теперь. Каждое лесное сообщество есть определенный индивидуум, внешние особенности которого не только дают возможность отличить один такой сложный организм от другого, но указывают также на внутреннее единство такого сообщества, Я хочу сказать, что такие лесные сообщества, отличимые Друг от друга с помощью вышеприведенных признаков, представляют собою не только внешнюю однородность, как об этом уже было сказано, но и внутреннее единство. Я хочу сказать этим, что каждому из них свойствен свой определенный биологический режим. За отличиями внешними идут различия внутренние. То, что наиболее существенно, а потому и наиболее Ценно в природе леса, это элемент взаимодействия, но характер и степень сложности его будут различны в зависимости от того, будет ли у нас насаждение чистое или смешанное, простая или сложная форма, густое или редкое стояние, большая или меньшая плотность населения и т. д. и т. д. Как всюду в биологии, так и в этой ее отрасли определенным формам соответствуют и определенные отправления.
Эти рассуждения и эти факты дают нам возможность связать объединяющие признаки с разъединяющими; определение леса, какое было дано и которое указывает на самый существенный признак – на элемент взаимодействия, – свойственно всякому лесу, но в разных его формах, т. е. в разных лесных сообществах эти элементы взаимодействия неодинаковы по степени, по характеру и т. д.
В лесах, подверженных хозяйству, приходится считаться с вопросом О происхождении насаждения; для лиственных, например, необходимо указание, произошло: ли оно из семян, или вегетативным путем из почек, развившихся на пне после вырубки деревьев, а то, может быть, из корневых отпрысков, если природа обладает таковыми, и т. п. Порослевое или вегетативное происхождение лесного сообщества, как мы увидим впоследствии, имеет большое биологическое значение в разнообразных направлениях. Свойства порослевого и семенного леса неодинаковы; знать происхождение леса необходимо не только в лесоводственных, но и в общих ботанико-географических целях. Есть целый ряд внешних признаков, по которым можно отличить порослевый лее от семенного; первый отличается более быстрым ростом в молодости при пневой поросли – групповым, или гнездовым, расположением деревьев. Вопрос о происхождении этого, лесного сообщества этими данными не ограничивается; во целому ряду признаков, которые можно отыскать в лесу, можно судить о том, какого рода было вмешательство, человека в его жизнь, под влиянием какой рубки он создался и т. д. Но входить во все эти вопросы мы сейчас не в состоянии.
Описав каждый ярус (если насаждение было сложное) в отношении состава, возраста, полноты, добротности, состояния, происхождения, надо спуститься еще ниже и описать подлесок, если таковой имеется; он описывается проще, чем ярусы: к «ему почти не применяется количественная характеристика [1 Современное лесное хозяйство СССР широко использует подлесочные растения как важную часть леса для удовлетворения различных потребностей народного хозяйства. Так, кустарник бересклет идет на выработку весьма ценного материала – гуттаперчи. В связи с этим в современном хозяйстве подлесок также часто учитывается весьма тщательно и точно. Ред.], ограничиваются указанием на состав пород в порядке их господства, на степень равномерности распределения, на густоту, различая только три степени, на среднюю высоту, а иногда и на возраст.
Теперь наступает очередь охарактеризовать подрост, если таковой имеется. Здесь опять-таки нужно прежде всего, конечно, указать на состав его, затем возраст и степень равномерности распределения его по площади, расположен ли он единично или группами, много ли его или мало, приурочен ли он к окнам в пологе насаждения (рис. 45) или встречается и непосредственно под кронами, степень его разновозрастности и, наконец, весьма важное качество его – степень его угнетенности. Целью его описания для лесоводов, является необходимость ответить на коренной вопрос: может ли он заменить собой материнское насаждение или нет, происходит ли смена пород и т. д. Для характеристики «благонадежности или неблагонадежности» подроста служат все перечисленные моменты как количественного, так и качественного характера.
После описания молодого поколения остается охарактеризовать живой и мертвый покров, а затем почву, которую лучше однако, описывать в связи с другими внешними признаками которые обычно разумеются под термином «местоположения или местопроизрастания».
Таким образом, мы завершили круг: начали с внешних признаков, перечислили внутренние и вновь вернулись к внешним, вот эти-то признаки в своей совокупности в различных своих сочетаниях и, наконец, каждый в отдельности могут служить достаточным основанием для расчленения леса на отдельные лесные сообщества, или для так называемого выдела насаждений.
Если раньше мы разумели под лесом, а стало быть и под любой его частью, под лесным сообществом, такую совокупность древесных растений, в которой обнаруживается взаимное влияние дерев друг на друга и на занятую почву и атмосферу, то теперь, в дополнение, мы можем сказать, что под сообществом, или насаждением, мы (разумеем часть леса, однородную в самой себе и отличающуюся какими-нибудь из перечисленных признаков от соседних. Это чисто эмпирическое определение дает возможность разъединить, расчленить лесной массив на отдельные сообщества. Признаки, которые выработало лесоводство, все такого рода, что или определяют общественную жизнь древесных растений, или ее характеризуют, представляя собою лишь внешнее выражение для той внутренней жизни, которая там протекает.
При той связности явлений, которую мы наблюдаем в лесу как между составляющими его живыми элементами – растениями, так и между ними и средою, можно заранее сказать, что не могут не иметь существенного значения такие обстоятельства, как простая или сложная форма леса, густое или редкое насаждение и т. д. и т. д. Если у нас есть сосновое насаждение определенного возраста, в определенных условиях местопроизрастания, то оно будет обладать определенными биологическими свойствами: в известной, свойственной данной пород и данным условиям местопроизрастания степени будет происходить там борьба за существование между его элементами; жизнь наличного поколения и его потомства будет находиться под определенным жизненными режимом в отношении количества света, которое будет пропускать полог, количества тепла, осадков, ветра и т. д.; будет там определенный почвенный покров; все друг с другом связано, одно обусловливает другое. Перед нами – целый сложный организм, между частями которого внутренняя и закономерная связь и который, как и подобает всякому организму, отличается известной устойчивостью: стоит в нем произвести какие-нибудь нарушения, природа их снова залечит, рано или поздно, во в нем все восстановится в соответствии с особенностями внешней среды и с внутренними свойствами составляющих его элементов. Но попробуем, например, искусственно ввести в состав такого соснового сообщества ель, хотя бы в виде подлеска. Влияние этого нового елового сообщества скажется прежде всего на ином почвенном режиме, так как почва не будет получать того количества осадков, которое получала раньше; подзолообразовательный процесс усилится, влажность почвы изменится – почва станет суше, холоднее; подстилка увеличится в своей мощности, может измениться в своих качествах, живой покров будет иной; до почвы станет меньше проникать света и тепла, и для самосева сосны, который радостно произрастал под материнским пологом, могут наступить тяжелые дни: значительная часть его может умереть от светового голодания.
В заключение этой главы необходимо сделать одно пояснение: очень часто употребляется термин насаждение, который должен был показаться совершенно чуждым; он употреблялся и будет дальше употребляться как синоним лесного сообщества или части леса однородной в себе и пр. и пр. Этот исконный лесоводственный термин приобрел уже гражданство и проник уже даже в ботаническую литературу. Неудачный характер его заключается, по-моему, в том, что с именем «насаждение» может связываться нечто насаженное, следовательно искусственно явившееся на свет божий, тогда как этот лесоводственный термин применяется ко всем участкам леса – безразлично, возникли ли они искусственно или представляют собою девственные леса. Французские лесоводы называют насаждение peuplement, немецкие – Holzbestand.
ПРИЧИНЫ НЕОДНОРОДНОСТИ ЛЕСА. ЛЕС КАК ЯВЛЕНИЕ ГЕОГРАФИЧЕСКОЕ
Почему же мало-мальски большой лесной массив не однороден? Потому, что он занимает большое пространство, а последнее в природе весьма редко бывает однородным. Лик земли в пределах пространства, которое занимают лесные массивы, обычно расчленен, обладает известным рельефом, пластикой. Это внешнее расчленение земной поверхности влечет за собой внутреннее расчленение как почвенной среды, с одной стороны, так прилегающих слоев атмосферы – с другой. Всем хорошо известно, что климатические условия северного и южного, восточного и западного склонов не одинаковы, что климат котловин, впадин и других отрицательных форм рельефа иной, чем холмов, бугров, разных выпуклин и других положительных форм рельефа. Все это справедливо как в отношении микро-, так и макрорельефа и для разных степеней между ними. Инсоляция, излучение, воздушные течения, осадки – все нормируется и определяется характером расчленения земной поверхности. То же самое справедливо и по отношению к распределению снега, стока поверхностных вод, их запруживанию и т. п. Мало того, это верно и по отношению к толще земли, по отношению к коре выветривания или еще иначе – к почве и грунту. Там, где поверхность земли совершенно ровная, там почвенный процесс, происходящий всегда под давлением местного климата, протекает так, а не иначе, но одинаково во всех частях; стоит только этой поверхности измениться, получить складки, углубления и т. д., как почвенный процесс и в силу дифференцировки и местного климата, и в силу иной циркуляции воды и других причин станет протекать неодинаково. Если рассматриваемое пространство велико, то, в дополнение к сказанному состав материнских пород, из которых образуется почва, может быть неодинаков. Но все вкратце указанные внешние условия имеют биологическую ценность по двум причинам: во-первых, потому, что сами растения не отличаются одинаковой потребностью в отношении света, тепла, влаги, зольных веществ, физических свойств почвы и т. д., а во-вторых, потому, что особенности внешней среды могут в известных пределах видоизменять наследственные биологические свойства растений, отражаясь на быстроте и предельности роста, величине растения, их формах, на плодоношении, на степени теневыносливости и т. д. Влияя на эти свойства, внешние условия среды, как это мы увидим впоследствии, сильно влияют на такие моменты общественной жизни растений, как степень энергии борьбы, за существование, степень изменения климатической обстановки под пологом, на особенности мертвого покрова, на появление и устойчивость самосева и т. д.
За расчленением земной поверхности идет расчленение почвенного покрова и прилегающих слоев атмосферы, а все взятое вместе влечет за собою и расчленение растительного покрова. В силу разных потребностей у разных видов прежде всего всегда заметно расчленение по составу: к более бедным или более сухим местам приурочиваются растения наименее требовательные, например сосна; на более богатых и плодородных почвах поселяются более требовательные элементы, например дуб с его спутниками; где избыток проточной влаги – растения, мирящиеся с этого рода условиями, например черная ольха вдоль уреза воды; по мере обогащения бедных песчаных почв другими элементами – на супесях, например, – к сосне могут примешиваться более требовательные представители древесной флоры, и с тем большею настойчивостью, чем более для них будут приемлемы условия местопроизрастания, так, прежние чистые сосновые насаждения будут постепенно превращаться в смешанные или сложные: из сосны и ели, из сосны; и дуба и т. д.
Многие изученные профили через некоторые лесные массивы свидетельствуют о том, как под влиянием иного состава почвенного покрова меняется состав лесных сообществ. Если мы первоначально не будем обращать внимания на некоторые детали в различиях и особенностях почвенного покрова, более или менее однородного в пределах целого района, то можем, как это показывает помещенный на странице профиль (рис. 46,), различить следующие типы рельефа, свойственные значительной части нашей лесостепной полосы. Правые высокие берега вдоль рек, покрытые моренными и лёссовидными суглинками, представляют собою обыкновенно сильно овражистую местность; за живым урезом воды начинается область поймы из аллювиальных наносов, затем песчаная надлуговая терраса, то более, то менее всхолмленная, и, наконец, опять более равнинная, так называемая переходная полоса к степи с супесчаными почвами. Этим четырем крупным подразделениям в местной геологии и рельефе соответствует и целая совокупность лесных сообществ: высокие правые берега покрыты дубовыми лесами, поймы – разнообразными лиственными лесами, вплоть до черноольховых трясин, надлуговые террасы – сосновыми борами, а переходные полосы к степи – сложными насаждениями из сосны в верхнем ярусе, дуба с некоторыми его спутниками – во втором и кустарникового подлеска, преимущественно и;; бересклета, – в третьем.
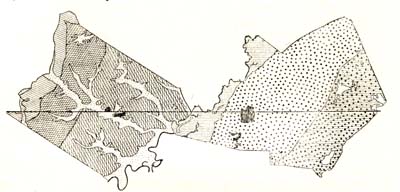
Рис. 46. Распределение насаждений в зависимости от рельефа и почвенно-грунтовых условий в пределах Воронежского окр.
Если мы теперь внимательно всмотримся в различия рельефа и почвенного покрова в пределах указанных четырех областей, то заметим следующие особенности, которые неизбежно влекут за собою и соответствующее, как увидим сейчас, различие в составе, росте и форме лесонасаждений. Подвигаясь от степи по направлению к реке, мы вначале встречаем обыкновенно более равнинную местность, выстланную темноцветными, темно-серыми или черноземновидными почвами; постепенный переход к более крутым поверхностям характеризуется и более светлыми – серыми – почвами, среди которых разбросаны отдельными оазисами так называемые солонцеватые суглинки и солонцы. В связи с таким расчленением почвенного покрова, как это и изображено на схеме (рис. 46), находятся и соответствующие различия в характере лесных сообществ; темноцветные и светлосерые почвы и соответственные им грунты населены, во-первых, сложными насаждениями, во-вторых, смешанными, в-третьих, лучшего, роста, чем лесные сообщества на солонцеватых местах; тогда как последние состоят из одноярусных, чистых и низкорослых, даже обычно суховершинных насаждений из дуба (рис. 47 и 48), лесные же сообщества из раньше наименованных состоят: в верхнем ярусе из дуба, ясеня, клена остролистного, во втором – из ильмовых, липы, полевого клена, в третьем – из того же паклена, неклена или татарского клена, лещины и т. п. В свою очередь, насаждения темносерых почв от насаждений на светлосерых почвах отличаются своим ростом или производительностью, характерным поведением ясеня, который на черноземновидных почвах лучше развит, перегоняет даже дуб и встречается притом в гораздо большем количестве; насаждения этой категории отличаются и более разнообразным составом и лучше развитыми кронами и, вероятно, лучшим плодоношением. Для иллюстрации того, как отражаются лучшие почвенные условия на росте лета, берутся модельные деревья, т. е. типичные, определенным методом выбранные, представители спелых насаждений всех трех категорий. Вычисленные объемы стволов этих моделей показали, что типичные представители черноземных почв превышают своих собратьев на солонцеватых почвах в 11—13 раз.
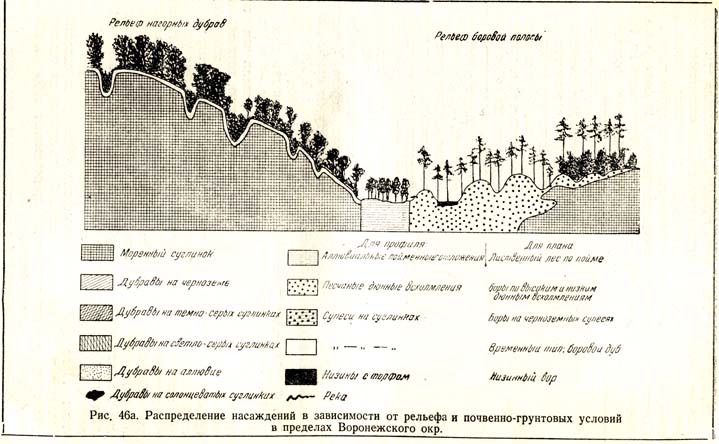
Если раньше различие в размерах дерева V класса по Крафту по сравнению с размерами той же породы на той же почве, но принадлежащей к господствующему классу — к I и II, (мы рассматривали как социальный гнет, то: в данном случае тоже большую, хотя все же меньшую разницу в объеме стволов, высоте и т. д. мы имеем право рассматривать как результат разных физико-географических условий, в частности как гнет почвенно-грунтовых условий. Таким образом, наш правый берег с его дубовыми лесами расчленился на три типа лесных сообществ, каждый из которых обусловлен различиями в почвенно-грунтовых условиях. Мы можем сказать, что наш правый берег состоит из насаждений трех категорий: 1) из дубрав на темно-серых почвах с определенными особенностями в отношении состава, формы, роста, плодоношения, а впоследствии увидим — ив поведении самосева; 2) из дубрав на светлосерых почвах с определенными различиями от первых и, наконец, 3) из дубрав на солонцеватых суглинках, где насаждения представляют собою почти чистые, одноярусные дубняки.
Но более внимательная экскурсия через подобные лесные массивы почти всегда покажет нам, что действительность отличается большей пестротой, чем данная схема. Среди аналогичных дубрав мы можем часто встретить осиновые насаждения на почвах, где должны были бы корениться дубовые леса и где они на самом деле и находятся, но где-нибудь поодаль от наблюдаемого места; то можем встретить такие же дубняки, но без подлеска, на почвах, где подлесок обыкновенно развивается; то замечаем насаждения двухъярусные, а не трехъярусные, где притом не только нижний, но и верхний ярус из дуба с его спутниками несет ясные следы своего порослевого происхождения Что же это такое? Если при одних и тех же почвенно-грунтовых условиях мы наблюдаем различия в характере насаждений притом устойчивые, а не случайные, то причина такого явления может скрываться только во влиянии такого важного фактора как вмешательство человека. Сплошные вырубки в дубовых лесах могут дать смену семенного поколения порослевым; если такая вырубка производится в возрасте, когда порослевая способность у дуба уже заглохла, то мы можем получить смену дуба его спутниками – грабом, буком, липой, в некотором случае – налет осины и впоследствии – осинники с подлеском из липы клена и т. д.; наконец, при неумеренной пастьбе скота легко уничтожается подлесок и т. д.