большой массивностью,
и мы вправе предполагать, что таранные кости
большой ширины принадлежат быкам. Существенно,
однако, что мы оказались не в состоянии
обнаружить ясной зависимости между длиной и
относительной шириной кости: очень широкие и
массивные экземпляры таранных костей имеют у
туров самую различную длину; то же может быть
сказано и о наиболее тонких, грацильных
экземплярах. Если мы сравним длину таранных
костей, отличающихся наибольшей массивностью
(ширина кости более 71% общей длины ее), с
остальными, подобной массивностью не
отличающимися, то получим следующие цифры:
Кол. |
Lim. |
M ± m |
|
Длина костей, имеющих ширину более 71% общей длины |
(n = 24) |
77 – 91 |
83,50±0,78 |
Длина костей, имеющих ширину менее 71% общей длины |
(n = 60) |
77 – 95 |
83,94±0,54 |
В обоих случаях, как видим,
средние арифметические значения общей длины
таранных костей совершенно сходны (M. diff. = 0,5) и о
каких-либо статистически достоверных различиях
не может быть и речи. Тонкие таранные кости имеют
в некоторых случаях даже большую длину, чем
массивные.
Все сказанное не означает,
разумеется, что половой диморфизм вовсе не
проявляется у туров в длине таранных костей.
Различия в этом отношении между быками и
коровами безусловно существуют, как имеются они
и у домашнего скота. Но различия эти, видимо,
совсем не абсолютны и не достигают таких
размеров, которые можно было бы предполагать в
данном случае, если основываться на мнении
Герре59 и Ленгеркена60, что у туров быки были
примерно на 50 см выше ростом в холке, чем коровы.
Еще более широкую изменчивость
общей длины таранных костей мы наблюдаем в
остатках из раскопок поселения Флорешты (таблица
7). В исследованной серии из 115 экземпляров,
найденных отчасти в слое культуры
линейно-ленточной керамики, но главным образом в
слое
58

Рис. 9. {th2-10-1.jpg} Таранные кости из
раскопок поселения Флорешты; слой культуры боян.
Обратить внимание на различия в размерах
культуры боян, длина колеблется
от 61 до 95 мм (рис. 9). Мы встречаем среди них,
следовательно, кости сравнительно небольших
размеров, вполне обычных у домашнего скота даже
очень поздних эпох, и наряду с ними экземпляры,
имеющие исключительно крупные размеры,
достигающие верхних пределов изменчивости,
отмеченных у аллювиальных туров. Первые образуют
левую часть вариационного ряда, вторые - его
правую часть. Если принять во внимание, что
минимальные размеры длины таранных костей у
туров (при доверительной вероятности 0,95)
составляют 74 мм, то тех же и более крупных
размеров достигают около 51% экземпляров из
раскопок поселения Флорешты и, следовательно,
лишь менее половины костей находится вне
пределов изменчивости, наблюдающейся у туров.
Совершенно аналогичную картину
широкой изменчивости мы обнаруживаем и при
изучении серии таранных костей из раскопок
поселений раннего триполья, к сожалению
немногочисленной и состоящей из 20 экземпляров
(таблица 7). Длина их колеблется от 62 до 96 мм, т. е. в
тех же пределах, которые были установлены в
остатках из поселения Флорешты. Почти две трети
экземпляров характеризуются размерами,
наблюдающимися у туров. Близкими в обоих случаях
оказываются и значения
59
средних арифметических;
статистически достоверные различия определенно
отсутствуют (М. diff. = 1,4).
Судя по данным, опубликованным
В. И. Бибиковой и А. И. Шевченко61, подобное же
явление имеет место и в остатках из раскопок
поселения позднеямной культуры Михайловка.
Длина таранной кости из упомянутого поселения
варьирует от 63 до 99 мм, причем, около 2/3 измеренных
экземпляров имеют длину более 74 мм, т. е.
свойственную одноименным костям туров. Сходство
наблюдается и в средних размерах костей (М. diff.
для серии из раскопок поселений Флорешты и
Михайловка всего 1,5).
Таким образом, анализируя
изменчивость общей длины таранных костей из
раскопок поселения Флорешты, поселений раннего
триполья и поселения Михайловка, мы во всех
случаях встречаемся с весьма сходной картиной. К
числу ее наиболее характерных особенностей
относится прежде всего чрезвычайно широкий
диапазон изменчивости. Во всех случаях мы
находим относительно мелкие экземпляры (длиной 61
- 63 мм), размеры которых свойственны, несомненно,
только домашнему скоту, и наряду с ними кости
чрезвычайно крупные, достигающие верхних
пределов изменчивости размеров, установленной у
туров. Именно по указанной причине диапазон
изменчивости кости оказывается в остатках из
упомянутых поселений гораздо более широким, чем
у туров или у какой-либо популяции
субфоссильного домашнего скота, взятых в
отдельности. Вряд ли отмеченное обстоятельство
может быть объяснено какими-либо иными
причинами, кроме признания, что в остатках из
раскопок всех этих поселений одновременно
встречаются кости диких и домашних особей.
Обращает на себя внимание и то,
что вариационные кривые изменчивости длины
таранной кости имеют в рассматриваемых случаях,
как и у туров, типично одновершинный характер, с
той лишь разницей, что вершина несколько
смещается влево и располагается в классах 70 - 74
или 74 - 78 мм. Следовательно, и здесь вариационная
кривая не дает каких-либо ясных указаний на
присутствие в остатках костей животных,
принадлежащих
60
группам, резко обособленным
друг от друга по размерам рассматриваемой кости
(этому должна была бы соответствовать по крайней
мере двухвершинная кривая). Вместе с тем
характерно, что положение вершин вариационных
кривых совпадает с нижними пределами
изменчивости длины кости у туров.
Все это заставляет думать, что
размеры длины таранных костей у туров и
домашнего скота из раскопок археологических
памятников, которые мы имели возможность
рассмотреть, несомненно трансгрессируют и что
трансгрессия эта весьма велика. Представляется
вполне вероятным, что ряд экземпляров, по длине
сходных с одноименными костями туров, в
действительности принадлежит уже домашним
особям, так же как турам принадлежат в отдельных
случаях кости длиной менее 76 - 77 мм, которые с
излишней категоричностью склонны относить
только к домашнему скоту.
К подобной же мысли приводит
изучение изменчивости длины таранных костей из
раскопок других археологических памятников. Так,
например, только в памятниках
восточноевропейского средневековья, где скот
отличается исключительно мелкими размерами62, мы
совершенно не встречаем костей, столь же крупных,
как у туров, т. е. длиной более 74 мм. (таблица 7).
Лишь изредка попадаются они в остатках из
раскопок городищ и поселений эпохи раннего
железа и поздней бронзы. Но в поселениях
неолитического и энеолитического времени они
вполне обычны, даже в тех случаях, когда нет
каких-либо указаний на присутствие в остатках
костей тура.
Особенно интересен в этом
отношении материал из раскопок энеолитических
поселений майкопской культуры на Северном
Кавказе. Изучение общего характера изменчивости
размеров таранных костей, в частности
компактность вариационного ряда, приводит к
убеждению, что кости из этих поселений
принадлежат только домашним особям, тогда как
остатки туров, видимо, отсутствуют совершенно.
Вариационные кривые изменчивости длины
рассматриваемых костей в остатках из раскопок
поселения Флорешты, а в значительной степени и
поселения Ми-
61
хайловка, обнаруживают большое
сходство с вариационной кривой изменчивости
костей из поселений майкопской культуры. Оно
проявляется в нижней границе изменчивости и в
величине модального класса. Разница лишь в том,
что в поселениях майкопской культуры
наблюдается менее широкий диапазон изменчивости
(длина костей не превышает 86 мм), с чем связано
заметное укорочение правого крыла ряда. Однако
примерно 40% экземпляров имеют длину более 74 мм, т.
е. достигают размеров свойственных турам.
Все это означает, по-видимому,
что в настоящее время мы можем считать вполне
определенно принадлежащими именно турам лишь те
экземпляры таранных костей, длина которых
превышает 86 мм и которые образуют крайне правые
классы вариационного ряда. Что же касается
экземпляров, длина которых находится в пределах
74 - 86 мм, то среди них в остатках из раскопок
памятников неолита, энеолита и раннего металла
могут встречаться кости как диких, так и домашних
особей - это зона трансгрессии.
Описанная изменчивость длины
таранных костей не представляет, однако, явления
универсального, свойственного остаткам из
раскопок памятников всех вообще энеолитических
культур Восточной Европы или хотя бы даже одной
только Молдавии. В этом с большой наглядностью
убеждает изучение материалов из раскопок
поселений культуры гумельница.
Очень широкий диапазон
изменчивости размеров таранных костей (59 - 93 мм)
наблюдается и в этих памятниках. Однако
вариационная кривая образует две ясно
выраженные вершины (рис. 10). Из 50 измеренных
костей 11, или 22%, отличаются очень крупными
размерами; длина их составляет от 83 до 93 мм.
Именно они и образуют правое, хорошо
обособленное крыло вариационного ряда; имеется
достаточно оснований считать эти крупные
экземпляры принадлежащими турам. Что же касается
остальной, гораздо более многочисленной (78%)
части серии, то к ней относятся не отличающиеся
крупными размерами экземпляры. Все они, за
исключением одной, имеющей длину 77 мм, находятся
вне пределов изменчивости, наблюдающейся у
туров. Мода выражена очень резко и расположена
ниже, чем в рассмотренных сериях, в классе 66 - 70 мм,
да и вся эта часть вариа-
62
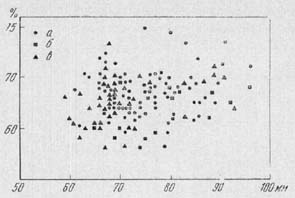
Рис. 10. {th2-11-1.jpg} График
корреляции общей длины и наибольшей ширины
таранных костей
На оси абсцисс - длина (в мм), на
оси ординат - ширина (в % к общей длине кости).
Условные обозначения: а - из раскопок поселения
Флорешты; б - из раскопок поселений раннего
триполья; в - из раскопок поселений культуры
гумельница
ционного ряда отличается
большой компактностью. Весьма показательно, что
в остатках из поселений культуры гумельница мы
констатируем резкую дифференциацию домашнего
скота и туров по длине таранных костей, чего
вовсе не наблюдалось при изучении материала из
раскопок Флорешты, поселений раннего триполья и
позднеямной культуры. Среднее арифметической
длины таранных костей из поселений культуры
гумельница (71,92±1,29) ниже, чем одноименных костей
из других неолитических памятников, хотя
статистически достоверных различий установить
не удается; они отчетливо выявляются при
сравнении с серией из поселения Михайловка (М. diff.
= 3,4). Если оставить в стороне кости длиной 83 - 93 мм,
которые мы считаем принадлежащими турам, то
длина таранных костей домашнего скота,
разводившегося племенами культуры гумельница,
составляет 59 - 77, в среднем 67,36±0,60 мм. Скот этот по
длине рассматриваемых костей уступает скоту
племен майкопской культуры (М. diff. = 4,4) и
приближается к скоту Восточной Европы в позднем
бронзовом веке, хотя и несколько крупнее его (М.
diff. = 4,4).
Нам остается еще коснуться
вопроса об изменчивости структурных
особенностей таранных костей (в частности,
63
Таблица 8.
Изменчивость индекса ширины
таранной кости скота (в % к общей длине её)
Скот из поселений
|
Вариационный ряд, мм |
Характеристики ряда |
||||||||||||
63,1 – 65,0 |
65,1 – 67,0 |
67,1 – 69,0 |
69,1 – 71,0 |
71,1 – 73,0 |
73,1 – 75,0 |
75,1 – 77,0 |
77,1 – 79,0 |
n |
Lim. |
М |
± m |
? |
C |
|
Тур |
2 |
13 |
28 |
17 |
11 |
8 |
4 |
1 |
84 |
63,6 – 78,4 |
69,58 |
0,33 |
3,04 |
4,4 |
Флорешты и раннее триполье |
2 |
23 |
30 |
27 |
10 |
3 |
1 |
– |
96 |
63,1 – 75,2 |
68,68 |
0,24 |
2,34 |
3,4 |
Культура гумельнила |
4 |
11 |
17 |
8 |
4 |
1 |
– |
– |
45 |
63,1 – 73,5 |
68,00 |
0,34 |
2,30 |
3,4 |
Таблицa 9.
Изменчивость общей длины
пяточной кости
Скот из поселений и памятников |
Вариационный ряд, мм |
Характеристики ряда |
||||||||||||||||||||
101,1 – 107,0 |
107,1 – 113,0 |
113,! – 119,0 |
119,1 – 125,0 |
125,1 – 131,0 |
131,1 – 137,0 |
137,1 – 143,0 |
143,1 – 149,0 |
149,1 – 155,0 |
155,1 – 161,0 |
161,1 – 167,0 |
167,1 – 173,0 |
173,1 – 179,0 |
179,1 – 185,0 |
185,1 – 191,0 |
n |
Lim. |
M |
±m |
? |
С |
||
Тур |
– |
– |
– |
– |
– |
– |
– |
– |
18 |
17 |
8 |
17 |
3 |
3 |
1 |
67 |
149 – 190 |
162,50 |
1,11 |
9,06 |
5,6 |
|
Флорешты |
– |
– |
– |
– |
8 |
12 |
9 |
5 |
10 |
3 |
1 |
4 |
4 |
– |
– |
56 |
126 – 179 |
145,76 |
1,93 |
14,46 |
9,9 |
|
Позднеямная культура |
– |
– |
– |
– |
– |
1 |
– |
3 |
4 |
– |
– |
– |
– |
1 |
1 |
10 |
132 – 186 |
155,00 |
4,22 |
13,34 |
8,6 |
|
Майкопская культура |
– |
– |
– |
– |
4 |
4 |
6 |
7 |
3 |
3 |
– |
– |
– |
– |
– |
27 |
128 – 159 |
142,22 |
1,76 |
9,12 |
6,4 |
|
Культура гумельницы |
– |
– |
– |
1 |
6 |
4 |
2 |
– |
1 |
4 |
– |
– |
– |
– |
– |
18 |
124 – 161 |
138,32 |
2.90 |
12,30 |
8,9 |
|
Позднебронзового века |
– |
– |
11 |
28 |
27 |
19 |
19 |
22 |
21 |
8 |
– |
– |
– |
– |
– |
150 |
115 – 163 |
136,10 |
1,00 |
12,18 |
8,9 |
|
Раннего железа |
14 |
29 |
77 |
130 |
108 |
89 |
40 |
26 |
8 |
– |
– |
– |
– |
– |
– |
521 |
102 – 155 |
126,32 |
0,44 |
10,08 |
8,0 |
|
Средневековья |
33 |
157 |
199 |
90 |
25 |
9 |
4 |
3 |
– |
– |
– |
– |
– |
– |
– |
520 |
101 – 148 |
117,0 |
0,31 |
6,96 |
6,0 |
|
отношения между наибольшей
шириной и наибольшей длиной, таблица 8).
Наиболее широко варьирует
рассматриваемый индекс (вычисленный на
основании приводимых Бекени измерений) у туров
(63,6 - 8,4%). Некоторое укорочение правого крыла
вариационного ряда наблюдается в остатках из
раскопок поселения Флорешты и поселений раннего
триполья, а в еще большей степени это имеет место
в серии из раскопок поселений культуры
гумельница. Нельзя не отметить, однако, что
модальные классы повсюду сходны. Вычисляя М. diff.
для величины индекса у туров и у животных из
раскопок поселений культуры гумельница, мы
устанавливаем, что различия статистически
достоверны (М. diff. = 3,4), т. е. ширина кости у туров
относительно больше. Однако приведенные в
таблице 8 данные показывают, насколько велика
трансгрессия рассматриваемого признака. В
подобных условиях использование его для
дифференциации костей диких и домашних особей
практически бесперспективно.
Существенно, что у животных из
раскопок энеолитических поселений, как и у туров,
мы оказываемся не в состоянии обнаружить
отчетливо выраженной зависимости между длиной
таранной кости и относительной шириной её (рис.
10). Очень массивные кости, свойственные, как можно
думать, быкам, равно как и тонкие, принадлежащие
коровам, имеют самую различную длину. Таким
образом, мы и в этом случае находим подтвержде-
64
ние предположению, что резкого
проявления полового диморфизма в длине таранных
костей не существует.
Перейдем теперь к рассмотрению
изменчивости общей длины пяточных костей
(таблица 9). По сравнению с таранными, они
представлены в исследованном материале менее
многочисленными сериями. Однако, и это очень
существенно, пяточные кости, у которых срастание
бугра с телом происходит примерно в трехлетнем
возрасте, являются тем самым материалом, хорошо
элиминированным по возрасту, чего нельзя сказать
о таранных костях. Изучение их по этой причине
имеет большой интерес и дает, как мы увидим далее,
весьма своеобразную картину изменчивости.
У аллювиальных туров
минимальные размеры общей длины пяточной кости
(рис. 11) определяются Бекени63 в 150, Бёсснеком64 - в
149, а К. Л. Паавером65 - в 161 мм. Суммируя измерения
этих костей, приведенные в упоминавшихся ранее
работах Бекени и Штампфли66, мы получаем весьма
своеобразный вариационный ряд. Вариационная
кривая в данном случае явственно двухвершинная,
причем правая вершина расположена в классе 167 - 173
мм, тогда как левая относится к крайнему ле-
65
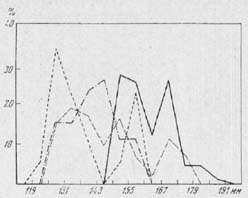
Рис. 11. {th2-12-1.jpg} График
распределения пяточных костей по общей длине.
На оси абцисс - длина (в мм.), на
оси ординат - количество экземпляров (в %).
Обозначения см. на рис. 8
вому классу ряда (143 - 155 мм.). Мы
встречаемся, следовательно, с явлением
асимметрии, выраженном даже ещё более резко, чем
у таранных костей. Уже одно это обстоятельство
может служить достаточно ясным указанием на
неполноту выборки, обусловленную искусственным
ограничением левого крыла вариационного ряда.
Даже при доверительной вероятности 0,95 у туров
должны встречаться пяточные кости длиной около
145 мм. Такие не отличающиеся крупными размерами
экземпляры упомянутые авторы относят всегда
только к домашним особям, чем порождается
неполнота и резкая асимметрия вариационного
ряда. Следует поэтому учитывать при изучении
костных остатков, что пяточные кости длиной от 145
до 150 мм. могут встречаться и у аллювиальных
туров. Что же касается причины, определяющей
двухвершинный характер вариационной кривой у
туров, то возникает мысль, что мы сталкиваемся в
данном случае с проявлением полового диморфизма
и что вершина в классе 167 - 173 мм. характеризует
кости быков, а вершина в классе 149 - 155 мм. - коров
туров.
66
Замечательное сходство в длине
пяточных костей обнаруживается между турами и
современным серым степным скотом67. У домашнего
скота (n = 7) она колеблется от 144 до 179, в среднем 162,4
мм, т. е. характеризуется цифрами, совершенно
типичными для аллювиальных туров.
Подтверждается, таким образом, картина, ранее
установленная при сравнении длины таранных
костей.
Интересно, что различие между
полами в длине пяточных костей выражено у серого
степного скота, насколько мы можем судить по
изученным нами семи экземплярам, совершенно
отчетливо. Представление об этом дают следующие
цифры:
Кол. |
мм |
||
Коровы |
(n = 3) |
150,3 |
(144 – 156) |
Быки |
(n = 4) |
171,5 |
(164 – 179) |
В среднем длина пяточной кости у
быков на 21 мм больше, чем у коров, т. е. различия,
учитывая общие размеры рассматриваемой кости,
очень велики. Интересно и то, что средние
величины длины кости у туров и быков серого
степного скота точно совпадают с вершинами
вариационной кривой изменчивости ее у туров.
Следует отметить, что различия
между полами у серого степного скота
обнаруживаются не только в абсолютных, но и в
относительных размерах пяточных костей.
Показательны в этом отношении размеры их,
выраженные в процентах от длины плюсневых
костей:
Кол. |
% |
||
Коровы |
(n = 3) |
58,7 |
(55,0 – 61,2) |
Быки |
(n = 4) |
65,0 |
(61,7 – 67,5) |
У быков, следовательно, длина
пяточных костей не только абсолютно, но и
относительно больше, чем у коров. Все это служит
дополнительным подтверждением справедливости
высказанного предположения относительно
природы наблюдающейся у туров двухвершинности
вариационной кривой длины пяточных костей.
68
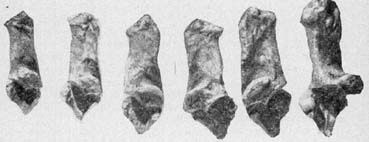
Рис 12. {th2-13-1.jpg} Пяточные кости из
раскопок поселения Флорешты: слой культуры боян.
Обратить внимание на различия в размерах.
С картиной очень сходной мы
встречаемся при изучении пяточных костей из
раскопок поселения Флорешты (рис 12). В имеющейся
серии из 56 экземпляров длина их варьирует от 126 до
179 мм. Это означает, что в ее составе имеются
кости, значительно уступающие по размерам
одноименным костям туров и несомненно
принадлежащие домашним особям, и наряду с ними
очень крупные, находящиеся в пределах
изменчивости, установленной для дикой формы;
последние даже многочисленны (около 40%
экземпляров). В этом отношении изменчивость
длины пяточных костей из раскопок Флорешты
повторяет ситуацию, ранее обнаруженную при
изучении таранных костей.
То же наблюдается и в остатках
из раскопок поселений раннего триполья, хотя, к
сожалению, из этой группы памятников мы
располагаем всего шестью экземплярами, у которых
tuber calcanei уже сросся с телом кости. Длина их
колеблется от 130 до 169 мм. Очень близкими
оказываются и средние размеры кости (145,7 мм в
серии из Флорешты и 144,3 мм в серии из поселений
раннего триполья).
Весьма интересно, что кривая
распределения длины пяточных костей из раскопок
упомянутых неолитических поселений Молдавии
имеет уже не две вершины, как это было
констатировано у туров, а явственно выраженные
три вершины. Две правые находятся в классах 167 -
69
173 и 149 - 155 мм, т. е. положение их
полностью совпадает с аналогичными вершинами
кривой у туров. Что же касается третьей, наиболее
крупной вершины, то она расположена в классе 125 -
131 мм, т. е. вне пределов изменчивости этой кости у
туров. Едва ли можно сомневаться, что она
образована вариантами, принадлежащими только
домашнему скоту. Столь же вероятно, что крайняя
правая вершина образована вариантами,
принадлежащими только турам. Что же касается
средней вершины, в классе 149 - 155 мм, то она может
иметь смешанное происхождение, включая как
экземпляры, принадлежащие коровам тура, так и
домашним быкам.
В пользу именно подобной
интерпретации этой средней вершины вариационной
кривой говорят не только результаты изучения
изменчивости пяточных костей у серого степного
скота, установившие существование значительных
различий по их размерам между быками и коровами.
По-видимому, в других случаях различия между
полами в этом отношении могут и не быть столь
значительными. Так, кривая распределения длины
пяточных костей у восточноевропейского скота
позднего бронзового века обнаруживает лишь
слабовыраженную дополнительную вершину, а
кривые распределения у скота раннего железного
века и средневековья явно одновершинные, хотя и с
сильно растянутым правым крылом. Возможно, что
вместе с уменьшением роста скота сокращается и
проявление полового деморфизма в размерах
костей.
Одновершинная кривая
наблюдается и у скота из поселений майкопской
культуры, хотя в этом случае она может найти себе
объяснение просто в ограниченности
исследованной серии.
Существенно, что у домашнего
скота пяточные кости могут иметь длину,
превышающую нижние пределы изменчивости ее у
туров. Об этом говорят, в частности, результаты
изучения их не только у серого степного скота, но
и у скота из раскопок поселений майкопской
культуры, в которых остатков туров не обнаружено
вообще. Пяточные кости у этого энеолитического
скота достигают 159 мм и около 25% общего количества
вариантов характеризуется размерами,
наблюдающимися у коров тура. Не только у столь
древнего, но и в исторически гораздо более
поздних популяциях скота приходится встре-
70
чать отдельные экземпляры
пяточный костей, по размерам не уступающих
одноименным костям туров, и лишь в памятниках
средневековья они почти совершенно отсутствуют.
Ясно, таким образом, что
говорить в подобных условиях о каких-либо очень
резких различиях между турами и домашним скотом
в длине пяточных костей решительно нет
оснований. Размеры пяточных костей у них,
несомненно, трансгрессируют, причем
трансгрессия в этом случае даже несколько шире,
чем у таранных костей. Можно считать, что
пяточные кости, имеющие длину менее 145 мм, всегда
принадлежат домашнему скоту, но совершенно нет
возможности утверждать, особенно при изучении
древних популяций, что каждая кость, превышающая
эти размеры, происходит обязательно от тура.
Не представляет поэтому простои
задачи интерпретация данных, полученных нами при
изучении пяточных костей из раскопок поселений
культуры гумельница. В пределах небольшой серии,
состоящей всего из 18 экземпляров, обнаруживаются
две резко обособленные группы. Одну из них
образуют кости длиной 124 - 139 мм, безусловно
принадлежащие только домашним особям (на их долю
приходится 13 экземпляров, или 72% измеренных).
Другую группу составляют кости длиной от 153 до 161
мм, которые могут происходить как от домашних
особей, так и от коров тура. Наблюдающийся разрыв
вариационной кривой в классе 143 - 149 мм имеет
скорее всего причиной ограниченность
исследованной серии. Учитывая общий характер
изменчивости пяточных костей, есть основания
предполагать, что среди костей длиной 153 - 161 мм
имеется хотя бы некоторое количество
экземпляров, принадлежащих домашним особям.
Характерно, что по величине среднего
арифметического кости из поселений культуры
гумельница лишь немного превышают кости скота
эпохи поздней бронзы.
Из длинных трубчатых костей
конечностей в исследованных нами костных
остатках из раскопок неолитических и
энеолитических памятников найдены хорошо
сохранившимися только отдельные экземпляры
пястей и плюсен. Хотя количество имеющихся целых
метаподий весьма ограничено, тем не менее
изучение их представляет большой интерес.
71
Общая длина пястных костей у
западноевропейских алллювиальных туров, по
сообщениям Бёсснека68 и Реквате69, составляет от 233
до 260 мм. Примерно такими же цифрами определяет ее
и Нобис70. В действительности же диапазон
изменчивости длины пястных костей несколько
шире, и это касается как верхних, так и нижних ее
пределов. Так, Бекени71 упоминает, что ему
приходилось встречать пясти туров длиной 219 мм,
причем одна из них происходила из раскопок
неолитического поселения, а другая была найдена
в памятнике римского времени. Известны и гораздо
более крупные, экземпляры; так, например, у
хранящегося в Геологическом музее АН УССР
скелета тура из Чаусова длина пясти достигает 269
мм. Следовательно, в настоящее время
изменчивость длины пясти у туров может быть
определена в пределах 219 - 269 мм. Впрочем, в
действительности размеры менее 230 и более 260 мм
встречаются относительно редко (таблица 10).
Интересно, что размеры длины
метаподий у коров и быков тура выражаются,
видимо, довольно близкими цифрами. По Бёсснеку,
например, длина пястей у коров 246 (240 - 253) мм, тогда
как у быков 250 (233 - 260) мм. По Нобису, длина костей у
первых 247 (243 - 253) мм, а у вторых 252 (240 - 258) мм.
Согласно измерениям, произведенным Бекени, длина
пясти у венгерских туров составляет у коров 241 (229
- 252) мм, а у быков 237 (219 - 252) мм. У туров из раскопок
неолитических поселений на территории Дании72
длина пясти у коров 246,8 (243 - 253) мм, а у быков 253,4 (248 -
258) мм. Как сообщает Штампфли73, длина пястных
костей коров тура из раскопок неолитического
поселения Зееберг-Бургешизее-Зюд 237 (235 - 240) мм,
тогда как у быков - 245 мм.
Суммируя имеющиеся в литературе
измерения пястных костей туров, а также
используя собственные наблюдения, можно
установить следующую характеристику
72
размеров и пропорций этой кости
у коров и быков (таблица 11). Мы видим, что хорошо
отличаясь гораздо большей массивностью, пясти
быков лишь немного превосходят длиной
одноименные кости коров.
Таким образом, половой
диморфизм, отчетливо проявляющийся в пропорциях
пястных костей (ширина эпифизов и диафиза
абсолютно и относительно больше, чем у коров),
лишь слабо выражен в общей длине рассматриваемой
кости.
Таблица 10.
Изменение общей длины пястных
костей
Скот |
Вариационный ряд, мм |
Характеристика ряда |
||||||||||||||||
150,1 – 160,0 |
160,1 – 170,0 |
170,1 – 180,0 |
180,1 – 190,0 |
190,1 – 200,0 |
200,1 – 210,0 |
210,1 – 220,0 |
220,1 – 230,0 |
230,1 – 240,0 |
240,1 – 250,0 |
250,1 – 260,0 |
260,1 – 270,0 |
n |
Lim |
M |
±m |
? |
C |
|
Тур из археологических памятников голоцена |
– |
– |
– |
– |
– |
– |
2 |
1 |
7 |
14 |
10 |
2 |
36 |
219 – 269 |
244,70 |
1,90 |
11,40 |
4,7 |
Из поселений позднего бронзового века |
– |
– |
1 |
15 |
30 |
19 |
11 |
1 |
– |
– |
– |
– |
77 |
177 – 223 |
198,50 |
1,17 |
10,30 |
5,2 |
Из памятников раннего железа |
5 |
15 |
106 |
137 |
80 |
36 |
4 |
– |
– |
– |
– |
– |
383 |
154 – 217 |
185,30 |
0,57 |
11,22 |
6,1 |
Из памятников средневековья |
15 |
90 |
291 |
205 |
34 |
4 |
– |
– |
– |
– |
– |
– |
639 |
152 – 205 |
177,60 |
0,35 |
8,77 |
4,9 |
Пястные кости,
достигающие хотя бы минимальных размеров
установленной у туров длины (219 мм), в остатках из
раскопок археологических памятников
восточноевропейского средневековья и раннего
железного века совершенно не встречаются
(таблица 10). Очень редкое явление представляют
они и в остатках из раскопок поселений эпохи
поздней бронзы, причем достигают именно лишь
минимальных размеров, известных у туров.
Таблица 11.
Остеометрическая
характеристика метаподий аллювиальных туров.
Признак |
Кость |
Корова |
Бык |
1. Длина кости, мм |
Пясть |
242 (229 – 253) |
247,6 (219 – 269) |
2. Ширина верхнего конца, мм |
Пясть |
70,6 (68 – 76) |
81,5 (68 – 90) |
3. Ширина нижнего конца, мм |
Пясть |
70,3 (65 – 80) |
82,5 (73 – 88) |
4. Ширина диафиза, мм |
Пясть |
39,1 (35 – 45) |
48,5 (42 – 53) |
|
Пясть |
29,2 (27,6 – 31,1) |
33,3 (30,5 – 35,7) |
|
Пясть |
28,8 (27,6 – 31,2) |
33,0 (30,2 – 35,2) |
|
Пясть |
16,1 (15,1 – 17,9) |
19,7 (18,1 – 21,3) |
5. Длина кости, мм |
Плюсна |
270 (265 – 285) |
280 (260 – 304) |
6. Ширина верхнего конца, мм |
Плюсна |
57,5 (57 – 58) |
68,8 (64 – 73) |
7. Ширина нижнего конца, мм |
Плюсна |
63,8 (62,0 – 65,5) |
74,7 (63 – 81) |
8. Ширина диафиза, мм |
Плюсна |
31,0 (31 – 32) |
34,9 (30,0 – 36,5) |
|
Плюсна |
21,1 (21,1 – 21,2) |
23,7 (22,6 – 25,4) |
|
Плюсна |
22,9 (22,1 – 23,8) |
26,6 (25,7 – 27,5) |
|
Плюсна |
11,1 (10,5 – 11,8) |
12,3 (11,7 – 13,5) |
74
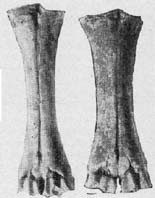
Рис. 13. {th2-14-1.jpg} Пяточные кости из
раскопок поселения Флорешты; слой культуры боян.
(слева - коровы, экз. № 2087; справа - быка, экз. № 1948).
Измерения см. в табл. 12
Кроме того, есть основания
думать, что эти наиболее крупные метаподии из
памятников поздней бронзы происходят от волов и,
следовательно, одноименные кости быков и коров
столь крупных размеров, как у тура, не достигали.
Длинные пястные кости, видимо,
более обычны у скота из более ранних памятников.
Во всяком случае Н. И. Бурчак-Абрамович74,
описавший остатки скота из раскопок курганных
погребений среднего бронзового века, приводит
измерения пястных костей, имеющих длину 203, 223 и 224
мм. В двух последних случаях пясти достигают
длины, уже встречающейся у туров, но и здесь дело
идет, впрочем, лишь о нижних пределах
изменчивости у дикой формы.