Хлоропласты (от греч. chlorós — зелёный и plastós — вылепленный, образованный), внутриклеточные органеллы растительной клетки — пластиды, в которых осуществляется фотосинтез. Окрашены в зелёный цвет благодаря присутствию в них основного пигмента фотосинтеза — хлорофилла. Основная функция Х., состоящая в улавливании и преобразовании световой энергии, нашла отражение и в особенностях их строения. У высших растений Х. — тельца линзообразной формы диаметром 3—10 мкм и толщиной 2—5 мкм, представляют собой систему белково-липидных мембран, погруженных в основное вещество — матрикс, или строму, и отграничены от цитоплазмы наружной мембраной (оболочкой). Внутренние мембраны образуют единую (непрерывную) пластинчатую, или ламеллярную, систему, состоящую из замкнутых уплощённых мешочков (цистерн) — т. н. тилакоидов, которые группируются по 10—30 (стопками) в граны (до 150 в Х.), соединяющиеся между собой крупными тилакоидами. При таком строении значительно увеличивается фотоактивная поверхность Х. и обеспечивается максимальное использование световой энергии. В мембране тилакоидов, состоящей из двух слоев белка, разделённых слоем липидов, осуществляется первичная световая стадия фотосинтеза, ведущая к образованию двух необходимых для ассимиляции CO2 соединений — восстановленного никотинамид-адениндинуклеотидфосфата (НАДФ×Н) и богатого энергией соединения аденозинтрифосфата (АТФ). Источником энергии для образования молекул АТФ является разность потенциалов, которая образуется на мембране в результате векторного (направленного) переноса заряда. Разделение заряда по обеим сторонам мембраны обеспечивается особым расположением компонентов электронно-транспортной цепи в мембране, перешнуровывающих её толщу. Благодаря мембранам, играющим роль «перегородок», осуществляется пространственное разобщение продуктов фотосинтеза, например O2 и восстановителей, без которых эти продукты взаимодействовали бы друг с другом. Наружная поверхность тилакоида покрыта частицами диаметром 14—15 нм, которые представляют собой «факторы сопряжения», участвуют в синтезе АТФ. В строме же сосредоточены ферменты фиксации CO2; (темновая стадия фотосинтеза).
У растений, способных к «кооперативному» фотосинтезу, существует 2 типа Х., различающихся по строению и функциям. Одни из них, находящиеся в клетках мезофилла, мелкие с гранами, другие, более крупные, содержатся в клетках обкладки проводящих сосудистых пучков, граны в них лишь зачаточные или совсем отсутствуют. В Х. второго типа функционирует фотосистема 1, которая образует АТФ в ходе циклического фосфорилирования, а НАДФ×Н — за счёт реакции декарбоксилирования яблочной кислоты. Х. клеток обкладки фиксируют CO2 на рибулозодифосфате, т. е. с помощью цикла Калвина, а Х. клеток мезофилла — на фосфоенолпирувате (путь Хетча — Слэка); т. о. взаимодействие Х. обоих типов обеспечивает высокую эффективность фотосинтеза у растений. В строму Х., наряду с ферментами фиксации CO2, включены нити ДНК, рибосомы, крахмальные зёрна, осмиофильные гранулы.
Наличие в Х. собственного генетического аппарата и специфической белоксинтезирующей системы обусловливает определённую, хотя и относительную, автономию Х. в клетке. При развитии и размножении растения в новых генерациях клеток Х. возникают только путём деления. Происхождение Х. связывают с симбиогенезом, полагая, что современные Х. — потомки сине-зелёных водорослей, вступившие в симбиоз с древними ядерными гетеротрофными клетками бесцветных водорослей или простейших.
Х. занимают 20—30% объёма растительной клетки. У водорослей, например хламидомонады, имеется один Х., в клетке высших растений содержится от 10 до 70 Х. Развиваются Х. из т. н. инициальных частиц, или пропластид, — небольших пузырьков, отделяющихся от ядра. В конце вегетации растения Х. в результате разрушения хлорофилла утрачивают зелёную окраску и превращаются в хромопласты. См. также Фотосинтез.
Лит.: Хлоропласты и митохондрии. Вопросы мембранной биологии, Сб., М., 1969; Лёви А., Сикевиц Ф., Структура и функция клетки, пер. с англ., М., 1971; Хит О., Фотосинтез, пер. с англ., М., 1972; Баславская С. С., Фотосинтез, М., 1974; Насыров Ю. С., Фотосинтез и генетика хлоропластов, М., 1975; Structure and function orchloroplasts, ed. М. Gibbs, B., 1971.
Р. М. Бекина.

Микрофотография хлоропласта.
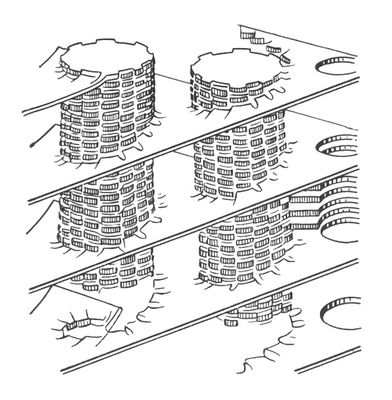
Модель пластинчатой (ламелярной) системы хлоропластов. Столбики — граны, образованные тилакоидами.