Трикарбоновых кислот цикл, цикл лимонной кислоты, цикл Кребса, широко представленный в организмах животных, растений и микробов путь окислительных превращений ди- и трикарбоновых кислот, образующихся в качестве промежуточных продуктов при распаде белков, жиров и углеводов. Открыт Х. Кребсом и У. Джонсоном (1937). Т. к. ц., локализованный в митохондриях, начинается с лимонной кислоты и заканчивается образованием щавелевоуксусной кислоты, CO2 и восстановлением коферментов дегидрогеназ: никотинамидадениндинуклеотида (НАД) и флавинадениндинуклеотида (ФАД). К субстратам Т. к. ц. относятся трикарбоновые кислоты — лимонная, цис-аконитовая, изолимонная, щавелевоянтарная и дикарбоновые кислоты — кетоглутаровая, янтарная, фумаровая, яблочная и щавелевоуксусная. К субстратам Т. к. ц. следует отнести и уксусную кислоту, которая в активной форме, то есть в виде ацетилкофермента А (ацетил-КоА), участвует в конденсации с щавелевоуксусной кислотой, приводящей к образованию лимонной кислоты. Именно ацетильный остаток, вошедший в структуру лимонной кислоты, подвергается окислению; атомы углерода окисляются до CO2, атомы водорода частично акцептируются коферментами дегидрогеназ, частично в протонированной форме переходят в раствор, то есть в окружающую среду.
Обычно указывают на пировиноградную кислоту (пируват), образующуюся при гликолизе в реакциях переаминирования и занимающую одно из центральных мест в перекрещивающихся путях обмена веществ, как на исходное соединение для образования ацетил-КоА. Действительно, под влиянием фермента сложной структуры — пируватдегидрогеназы — осуществляется окисление пирувата с образованием CO2 (первое декарбоксилирование), ацетил-КоА и происходит восстановление НАД (см. схему). Однако окисление пирувата далеко не единственный путь образования ацетил-КоА, который является характерным продуктом митохондриального окисления жирных кислот (фермент тиолаза), а также реакции обратной конденсации при образовании лимонной кислоты и др. Все ферменты, участвующие в реакциях Т. к. ц., локализованы в митохондриях, причём большинство из них прочно связаны с мембранными структурами.
Образование лимонной кислоты, с превращения которой и начинается собственно Т. к. ц., является реакцией эндергонической, и её реализация возможна благодаря использованию богатой энергией связи ацетильного остагка с KoA [СН3(О) С~SKoA]. Далее следует изомеризация лимонной кислоты в изолимонную через промежуточную стадию образования цис-аконитовой кислоты. Продуктом дальнейшего превращения изолимонной кислоты под влиянием соответствующей дегидрогеназы является, по-видимому, щавелевоянтарная кислота, декарбоксилирование которой (вторая молекула CO2) приводит к a-кетоглутаровой кислоте. Кетоглутаратдегидрогеназа по ряду характеристик (высокая молекулярная масса, сложная многокомпонентная структура, ступенчатые реакции, частично те же коферменты и т.д.) напоминает действие пируватдегидрогеназы. Продуктами реакции являются CO2 (третье декарбоксилирование), НАДН×Н+ и сукцинил-КоА. На этой стадии включается сукцинил-КоА-синтетаза, катализирующая обратимую реакцию образования свободного сукцината: Сукцинил-КоА + Рнеорг. + ГДФ Û Сукцинат + KoA + ГТФ. При этой реакции осуществляется так называемое субстратное фосфорилирование, то есть образование богатого энергией гуанозинтрифосфата (ГТФ) или аденозинтрифосфата (АТФ) за счёт гуанозиндифосфата (ГДФ) и минерального фосфата (Р) с использованием энергии сукцинил-КоА. После образования сукцината вступает в действие сукцинатдегидрогеназа — флавопротеид, приводящий к фумаровой кислоте. Фумараза обеспечивает равновесие между фумаровой кислотой и яблочной, а дегидрогеназа яблочной кислоты (кофермент — НАД+) приводит к завершению Т. к. ц., то есть к образованию щавелевоуксусной кислоты. На этой стадии повторяется реакция конденсации (конденсирующий фермент) между щавелевоуксусной кислотой и ацетил-КоА, приводящая к образованию лимонной кислоты.
Энергетическая эффективность рассмотренных процессов невелика. Однако образующиеся при окислении пирувата и последующих реакциях Т. к. ц. 4 моля НАДН, 1 моль ФАДН2 и 3 моля CO2 являются важными продуктами окислительных превращений. Особенно это касается восстановленных форм НАД и ФАД. Дальнейшее их окисление осуществляется ферментами дыхательной цепи и сопряжено с фосфорилированием, то есть образованием АТФ за счёт этерификации минерального фосфата (см. Окислительное фосфорилирование). На каждую полностью окисленную до CO2 и H2O молекулу пирувата приходится образование не менее 15 богатых энергией фосфатных связей. Процесс окисления НАДН и ФАДН2 ферментами дыхательной цепи энергетически весьма эффективен, происходит с использованием кислорода воздуха, приводит к образованию воды и служит основным источником энергетических ресурсов клетки. Однако в его непосредственной реализации ферменты Т. к. ц. не участвуют. См. также Окисление биологическое, Тканевое дыхание.
Лит.: Кребс Г., Корнберг Г., Превращения энергии в живых системах, пер. с англ., М., 1959; Филиппович Ю. Б., Основы биохимии, М., 1969; Ленинджер А., Биохимия, пер. с англ., М., 1974, гл. 16.
С. Е. Северин.
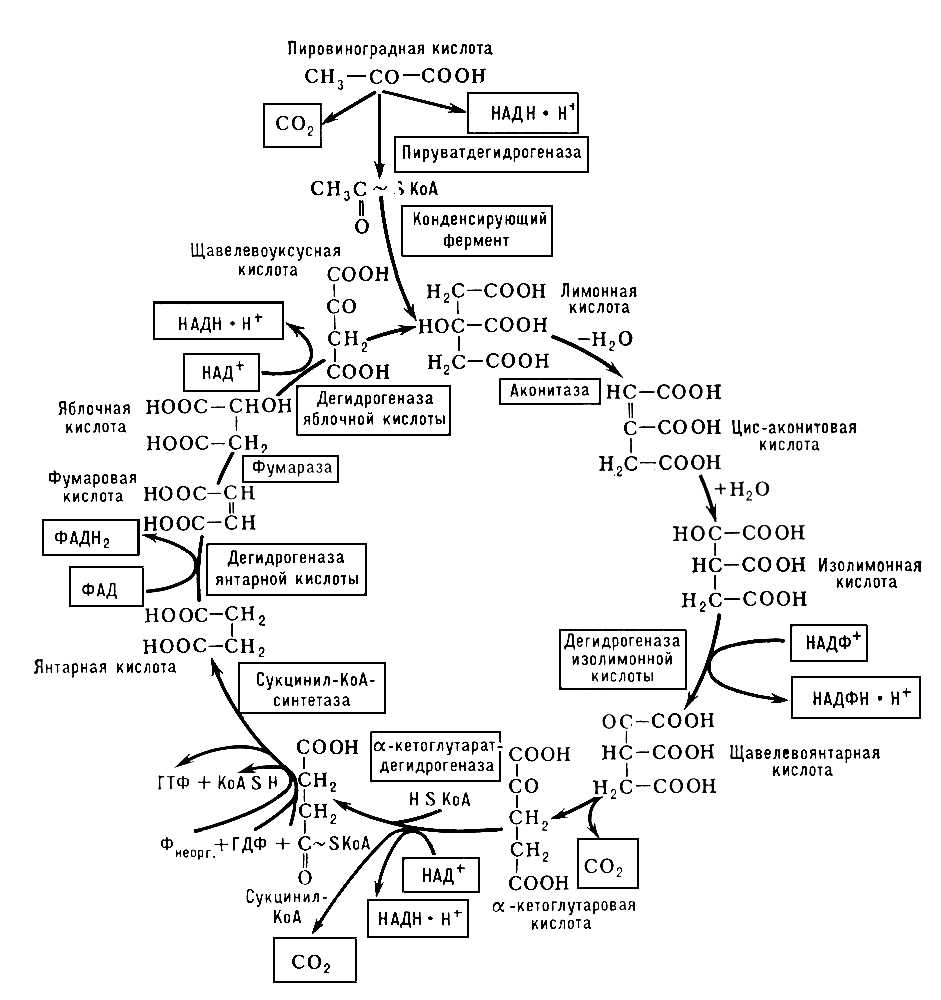
Схема цикла трикарбоновых кислот. В рамках — ферменты и окислённые формы коферментов, в двойных рамках — восстановленные коферменты и CO2.