Оплодотворение у pacтений
состоит в слиянии двух половых клеток — мужской и женской. Произошедшая через такое слияние клетка производит новое растение. При неизменности сути, процесс оплодотворения протекает различно у разных растений; равным образом весьма различно строение мужских и женских половых клеток, из которых первые оплодотворяют и играют в процессе активную роль, вторые являются оплодотворяемыми, и их участие более пассивное. Во многих случаях процесс О. в растительном царстве с его внешней, точнее морфологической стороны прослежен очень точно и обстоятельно; гораздо менее известны внутренние явления, совершающиеся при этом — физиология оплодотворения, хотя и в этом направлении сделаны уже интересные и удачные попытки (см. книгу Клебса, ниже). Несомненно, что в привлечении мужских элементов к женским видную роль играют так назыв. хемотактические и хемотропические явления, примеры чему будут указаны ниже. Во всяком случае для экспериментальных исследований тут открывается обширное и многообещающее поле. В противоположность низшим, споровым растениям, с их весьма разнообразными формами полового акта, у всех высших цветковых растений О. протекает довольно однообразно. В различных статьях, посвященных описанию отдельных групп низших растений (напр., Водоросли, Зеленые водоросли, Грибы), было сообщено и относительно процесса оплодотворения у этих растений; здесь остается, дополнив эти данные существенными подробностями, систематизировать различные формы О. и описать О. у высших, цветковых (иначе семянных) растений. Прежде всего необходимо, однако, указать, что процесс О. не найден до сих пор у довольно многих низших растений, из самых различных групп и отрядов, напр. его нет у бактерий и родственных им сине-зеленых водорослей (иначе водорослей-дробянок) — организмов, весьма низко стоящих по своему строению и развитию; нет его также у многих грибов — у базидиальных (к которым относятся между прочим наши съедобные грибы и "поганки"), далее у многих сумчатых, вероятно, у большинства их. У ржавчинных и головневых грибков существование О. пока недостаточно доказано. Предполагают, что грибы эти когда-то обладали оплодотворением, но постепенно утратили его, сохранив бесполые способы размножения. Их называют поэтому апогамными, в отличие от форм агамных, которые никогда, по-видимому, не имели оплодотворения. К таким агамным растениям принадлежат вышеупомянутые бактерии и сине-зеленые водоросли [В ст. Апогамия это явление неверно определено]. Превосходные случаи несомненной апогамии представляют некоторые папоротники, напр. Pteris cretica, y которого молодые растеньица вырастают всегда бесполым путем и как раз из тех мест заростка, где должны были находиться женские половые органы. Другой папоротник Aspidium filix mas. var. cristatum стал апогамным лишь под влиянием культуры. — Клетки, сливающиеся при акте О., получили название гамет. Они либо обладают самостоятельным (активным) движением, либо нет. В первом случае их называют плано-, во втором аплано-гаметами. Если сливающиеся гаметы одинакового строения, т. е. нельзя заметить разницы между мужской и женской гаметами, то слияние их и процесс оплодотворения называют изогамным, изогамией, при существовании различий — гетерогамным, гетерогамией. Между той и другой формой О. существуют формы промежуточные, переходные. Далее, при изогамии может происходить слияние (копуляция) двух плано- или же двух аплано-гамет. При гетерогамии бывает слияние плано- с апланогаметой или же двух апланогамет.
Изогамия. Копуляция планогамет. Эта форма О. встречается у многих зеленых водорослей, например у Ulothrix, у Botrydium (обе водоросли изображены на таблице при ст. Водоросли; см. также ст. Зеленые водоросли). У той и у другой водоросли образуются два сорта зооспор, одни для бесполого размножения, другие для полового. Половые зооспоры-гаметы маленькие, грушевидной формы, каждая с 2 ресничками. Из каждой клетки Ulothr i x их образуется 16—32, а у Botrydium помногу. Гаметы копулируют, сливаясь по две, реже по 3, а иногда даже по 4 (см. фиг. 5 b, с, 12 с. тбл. Водоросли). При этом они сцепляются сначала бесцветными носиками, на которых сидят реснички, потом поворачиваются, прикладываются друг к другу бочком, наконец постепенно сливаются (все время продолжая двигаться) в крупную зооспору уже с 4 ресничками. Зооспора скоро останавливается, теряет реснички и превращается в неподвижный шарик, окружающийся оболочкой (фиг. 5 d, е). Продукт копуляции называют зигоспорой или короче зиготой, также изоспорой (как продукт изогамии). Зигоспора превращается в новое растеньице непосредственно или производя предварительно зооспоры (фиг. 5 f; g), из которых каждая вырастает в новую водоросль. Интересно, что гаметы у Ulothrix копулируют только тогда, когда происходят из 2 различных нитей. Кроме того, по Клебсу, копуляция гамет происходит только при температуре ниже 25° Ц., уже при 26° — 27° Ц. копуляции не бывает, а зоогонидии прямо останавливаются, округляются и превращаются в споры, прорастающие в новую водоросль. Впервые копуляция была подмечена (Прингсгейм, 1869) у Pandorina morum, маленькой шарообразной водоросли, представляющей из себя колонию из 16 клеток, похожих на крупные зооспоры, каждая клетка с 2 ресничками. Клетки толстыми концами обращены к центру колонии, а узкими носиками наружу. Колония окружена слизистой оболочкой, через которую высовываются наружу 32 реснички, служащие для передвижения всей колонии. При бесполом размножении каждая из 16 клеток распадается повторным делением на 16 маленьких клеточек, образующих новую колонию пандорины. При половом размножении клетки колонии также распадаются на маленькие клеточки, но последние не слагаются в колонию, а выплывают наружу в виде мелких двуресничатых зооспор — гамет. Гаметы копулируют вышеописанным способом и образуют зиготу, из которой выходит через некоторое время (так наз. "период покоя") одна крупная зооспора, которая затем распадается на 16 маленьких клеточек, слагающихся в новую колонию. Примечательно, что у пандорины гаметы не одинаковой величины. Смешав гаметы из 2 различных колоний, видим, что копулируют гаметы как одинаковой величины (т. е. мелкая с мелкой, крупная с крупной), так и разной (мелкая с крупной), и последнее бывает даже чаще. Очевидно, пандорина обнаруживает стремление перейти от изогамии к гетерогамному оплодотворению.
Изогамия. Копуляция апланогамет. Этот тип встречается как у водорослей, так и у грибов. Отличный, давно известный пример такого рода полового акта дает нам спирогира (Spirogyra) и близкие к ней водоросли — их соединяют в группу конъюгат (Conjugatae) именно на основании особенностей их полового процесса (конъюгация то же, что копуляция). Среди грибов эту форму О. находим у мукоров (Mucor) и их сродичей, представителей целой группы, называемой Zygomycetes. Спирогира — весьма обыкновенная пресноводная водоросль, ее зеленые ниточки заметны и простым глазом. Под микроскопом в клетках бросаются в глаза одна или несколько изящных ленточек, спирально завитых и зазубренных по краям (фиг. 39 тбл при ст. Водоросли; см. еще ст. Зеленые водоросли), — это хроматофор, их называют также хлорофильными лентами. При копуляции содержимое одной клетки сливается с содержимым другой, причем сначала клетки двух рядом лежащих нитей дают сбоку выростки — отроги, растущие навстречу друг другу (фиг. 39); скоро отроги эти сталкиваются, разделяющая их перегородка растворяется и таким способом образуется канал, через который все содержимое одной клетки переливается в другую (фиг. 40). То содержимое, которое переливается, считается мужской гаметой, которое остается на месте — женской. Бывает, что сливается содержимое не супротивно-лежащих клеток, а соседних в одной и той же нити, в таком случае копуляционный канал огибает поперечную перегородку, разделяющую эти клетки. Во всяком случае продукт копуляции облекается собственной толстой оболочкой и превращается в зиготу (фиг. 40 b, с.), из которой весной вырастает новая ниточка спирогиры. Особенно примечательны с точки зрения изучения процесса О. явления, происходящие внутри зиготы (Хмелевский). Сначала в зиготе две хлорофильных ленты — одна женская, другая, пришедшая с мужской гаметой. Мужская лента начинает съеживаться, желтеть и постепенно разрушается и остается одна женская, так что потомство получает хроматофоры от одной матери; подобное мы встречаем и у высших цветковых растений (см. ниже). Что касается до клеточных ядер, то мужское сливается с женским, но дальше ядро снова делится на 2 новых, а эти еще раз, так что получается 4 ядра. Два из них скоро сближаются и сливаются в одно, а два других распадаются и исчезают, — в зиготе опять находится только одно ядро. Сходные явления замечены Клебаном у клостерия (Closterium), водоросли, родственной спирогире по развитию, но одноклетной и имеющей вид маленького, слабо изогнутого серпа. У клостерия мужское и женское ядра также сливаются в зиготе, но позже, именно только весной при прорастании зиготы. Произошедшее через слияние ядро затем делится пополам и каждая половина делится еще раз. В то же время распадается и все содержимое зиготы пополам, так что образуются 2 клостерия и в каждом по 2 ядра. Но главный интерес в том, что и здесь сохраняется только одно ядро, более крупное, а другое, поменьше, разрушается и скоро совсем исчезает. Не выходя из пределов группы Conjugatae, мы встречаемся с формами, у которых разница между мужской и женской гаметами выражена в большей или наоборот в меньшей степени, нежели у спирогиры. Так у Sirogonium клетки с мужскими гаметами меньше чем с женскими, ясно, что это уже стремление перейти к гетерогамии. Наоборот у Меsocarpus обе гаметы направляются навстречу друг другу и сливаются на полпути посередине копуляционного канала, образуя здесь зиготу. Вследствие этого у Mesocarpus нельзя различить мужскую от женской гамету и мы имеем случай строгой изогамии. В равной степени изогамны десмидиевые водоросли (Desmidiaceae), к числу которых принадлежит и вышеупомянутый клостерий. Подобно тому как у Mesocarpus совершается копуляция у плесневого грибка — мукора, Mucor (см. статью Грибы и приложенную к ней табл. II). Зигоспора мукора, прорастая, дает росток, на верхушке которого обыкновенно образуется спорангий (табл. II, фиг. 2). В некоторых случаях копуляционные отроги мицелия у мукора, не успев слиться, приносят тем не менее на концах своих по споре и притом совершенно похожей на зигоспору. Это случай утраты полового акта — апогамии. Такие споры называют азигоспорами.
Гетерогамия. Слияние мужской планогаметы с женской апланогаметой. Весьма распространенная форма полового акта, при которой мужские гаметы всегда активно подвижны; они носят название живчиков, сперматозоидов или антерозоидов и образуются в специальных органах антеридиях; женские гаметы неподвижны, их называют оосферами или яйцами, так как они соответствуют вполне яйцам животных, они формируются в оогониях или, у более высоко организованных споровых растений — в архегониях. Эту форму полового процесса, называемую оогамией, хорошо можно наблюдать у двух часто встречающихся у нас пресноводных водорослей — эдогония (Oedogonium) и вошерии (Vaucheria; см. ст. Зеленые водоросли и тбл. при ст. Водоросли). У эдогониев образуется в оогонии одна шарообразная оосфера или яйцо темно-зеленого цвета с бесцветным воспринимающим пятном на вершине (фиг. 7). Сперматозоиды имеют вид маленьких зооспор с венцом ресничек вокруг бесцветного носика (фиг. 9, b). Проникнув через отверстие в оболочке оогония, сперматозоид утыкается носиком в воспринимающее пятно яйца (фиг. 7) и скоро совсем сливается с яйцом. Оплодотворенное яйцо сейчас же окружается оболочкой, вследствие чего становится невозможным проникновение в него других сперматозоидов, и превращается в ооспору. Применяя метод окраски Клебана, оказывается, что протоплазма сперматозоида сливается с протоплазмой яйца, а его ядро постепенно приближается к ядру яйца — сначала оно значительно меньше ядра яйца и более компактное, но по пути несколько увеличивается и как бы разрыхляется — скоро оно достигает ядра яйца и мало-помалу с ним совершенно сливается. После периода покоя (у одних видов более короткий, у других более продолжительный — целую зиму, например) содержимое ооспоры распадается на 4 крупных зоооспоры, из коих каждая вырастает в новую ниточку эдогония. У вошерии оплодотворение происходит обыкновенно ночью и, если хотят следить за ним днем, то нужно водоросль с вечера поместить на лед. Перед О. у вошерии небольшая часть содержимого оогония выбрасывается наружу в виде бесцветной слизистой капли, а остальное содержимое формируется в яйцо. Уже через несколько минут после погружения очень маленького сперматозоида в воспринимающее пятно яйца, оплодотворенное яйцо окружается тонкой оболочкой, а спустя несколько часов исчезает и воспринимающее пятно и ооспора становится равномерно зеленого цвета. Зрелая ооспора красно-бурого цвета, покрыта толстой оболочкой и переполнена крупными каплями масла, служащего запасным питательным веществом для будущей молодой вошерии, которая вырастет из ооспоры. К тому же отделу Зеленых водорослей (Chlorophyceae), к которому принадлежат только что упомянутые водоросли, относится и вольвокс (Volvox), подобно пандорине — шарообразная подвижная колония, состоящая из гораздо большего числа клеточек (см. тбл. при ст. Водоросли, фиг. 4). Шарики вольвокса заметны и простым глазом. При половом размножении только часть клеток превращается в половые органы; при этом одни из них разрастаются в грушевидные оогонии с одной круглой оосферой внутри, а другие превращаются в антеридии,и внутри которых образуется много мелких булавовидных живчиков (сперматозоидов) с 2 ресничками (фиг. 4). В массе живчики желтого цвета, оосферы — темно-зеленые. Оплодотворенная живчиком оосфера облекается оболочкой и превращается в ооспору. Таким образом, у вольвокса типичная оогамия. Но существует водоросль, родственная пандорине и вольвоксу — Eudor i na elegans, у которой О. происходит в форме, средней между изогамией и оогамией. Ко времени полового размножения колонии Eudorina дифференцируется на женские и мужские. Каждая из клеток женской колонии превращается в шаровидное яйцо, каждая клетка мужской колонии делится повторно и превращается в целый пучок маленьких живчиков, похожих на живчики вольвокса. Высвободившись из колонии и плавая в воде, пучки живчиков наталкиваются на женские колонии и распадаются. Тогда живчики добираются до яиц и один из них сливается с яйцом. Оплодотворенное яйцо превращается, как обыкновенно, в ооспору. Подобные переходные по своему О. водоросли существуют также и между так называемыми темноцветными или бурыми водорослями, — см. ниже. У фукусов (см. это слово и таблицу при ст. Водоросли, фиг. 49, 50), крупных морских водорослей коричневого цвета различных оттенков, половое размножение (бесполого у них совсем не известно) происходит в форме типичной оогамии. Оогонии и антеридии образуются у них в особых ямках — вместилищах, так называемых conceptacula (см. табл. при ст. Водоросли, фиг. 22). В оогонии образуется не одно, а 8 круглых яиц (фиг. 23, 24). Впрочем, у других бурых родственных фукусу водорослей образуется яиц меньше — 4—2 или даже всего одно, как у атлантической Himan t halia lorea. Живчики формируются в мешочках-антеридиях помногу, они очень мелки и имеют 2 реснички, отходящие сбоку (фиг. 25—27). В море во время отлива зрелые яйца выталкиваются из оогония на свободу и скопляются у отверстия conceptaculum; к этому же времени и антеридии отрываются от несущих их веточек и скопляются там же. С наступлением прилива антеридии открываются и выпускают живчиков, устремляющихся к яйцам и начинающих так быстро вертеться около них, что огромные, по сравнению с ними, яйца начинают вихрем кружиться в воде (фиг. 28). В конце концов один из живчиков проникает в яйцо и оплодотворяет его, после чего оно покрывается оболочкой и превращается в ооспору, из которой вырастает новый фукус. Проще организованные бурые водоросли и размножаются проще. Из них морская бурая водоросль — кутлерия (Cutleria) производит подвижные двуресничатые гаметы, но различной величины: одни покрупнее — они образуются по одной в клетке, другие помельче — их образуется в клетке по несколько. Поплавав немного, крупная гамета останавливается, втягивает реснички, округляется и превращается в яйцо, при этом ее бесцветный носик переходит в воспринимающее пятно яйца. Тогда к ней подплывают мелкие гаметы, одна из которых утыкается носиком в воспринимающее пятно и скоро совсем сливается с яйцом — очевидно, это мужская гамета, вполне соответствующая сперматозоиду фукусов. Таким образом, на наших глазах происходит превращение женской планогаметы в неподвижное яйцо. Из сказанного выше можно заключить, что гетерогамия и специально оогамия развилась из более простой формы оплодотворения — из изогамии. Очевидно, что изогамные водоросли — более древние, половые же различия возникли лишь со временем и постепенно. Для всех более высоко организованных споровых растений (лучиц, мхов, папоротников, хвощей, плаунов) оогамия оказалась наиболее удобной формой полового акта; она стала у них единственной формой О. и вполне закрепилась. У всех у них мужские элементы — сперматозоиды различного строения, образующиеся в антеридиях. Яйца формируются, однако, не в одноклетных оогониях, как у водорослей и грибов, а в органах более сложного устройства, в так наз. архегониях. Что касается до устройства половых органов и внешней стороны процесса О., то об этом сказано в специальных статьях, трактующих об этих растениях; см. также Заросток. При О. сперматозоиды устремляются во множестве внутрь архегония, но только один из них сливается с яйцом. Оплодотворенное яйцо покрывается оболочкой, потом делится и производит новое поколение. Исследования Пфеффера показали, что у мхов и папоротников живчики привлекаются внутрь архегония различными веществами, выделяющимися из архегония вместе со слизью (хемотаксис живчиков). Вещества, действующие специфически на живчиков, различны; у папоротников, напр., яблочная кислота (ее содержится в слизи архегония около 0,3 %), у лиственных мхов — тростниковый сахар, у печеночного мха Marchantia — пока неизвестное вещество. Что дело тут именно в химич. раздражении живчиков, показывают опыты Пфеффера с искусственным вовлечением живчиков в капиллярные трубочки. Если запаянную на одном конце стеклянную капиллярную трубочку наполнить под колоколом воздушного насоса очень слабым (0,01—0,1 %) раствором какой-нибудь соли яблочной кислоты и затем погрузить открытым концом в каплю воды с живчиками папоротника, то последние устремятся внутрь трубочки. Для опыта с живчиками мха заменяют яблочную кислоту тростниковым сахаром.
Гетерогамия. Слияние двух апланогамет. Такого рода гетерогамия встречается у грибов, у водорослей и почти у всех цветковых растений. Среди грибов ее находим у представителей двух семейств: у пероноспоровых (Peronosporeae) и у сапролегниевых (Saprolegnieae). Женские половые органы пероноспоры (Peronospora) образуются на концах ветвей в виде шарообразных органов, наполненных густым содержимым. Это — оогонии; содержимое их дифференцируется на две части: центральную густозернистую, гоноплазму, превращающуюся в одну оосферу или яйцо, и периферическую, не столь зернистую, не принимающую участия в образовании яйца и наз. периплазмой. Мужские органы залагаются под оогонием в виде боковых веточек гифы, несущей оогонии. Веточки, нарастая, загибаются и прикладываются к оогонию, в то же время концы их вздуваются булавой и превращаются в антеридии. В этих антеридиях, однако, никогда не образуется живчиков. О. происходит иначе. Антеридий пускает отрог, который пробуравливает оболочку оогония и доходит до самого яйца, тогда верхушка отрога вскрывается и часть содержимого антеридия — зернистая гоноплазма — переливается к яйцу и сливается с ним. В антеридии остается, как и в оогонии, лишь постенный слой плазмы — периплазма. Оплодотворенное яйцо покрывается сначала тонкой оболочкой, которая постепенно утолщается, потребляя на образование своего наружного слоя, так наз. эписпория, периплазму. При прорастании ооспора дает прямо нить мицелия или же предварительно производит зооспоры. У сапролегний половые органы устроены так же, как и у пероноспоры (см. табл. при ст. Грибы, фиг. 10), но в оогониях не бывает периплазмы, а все содержимое оогония распадается на несколько яиц (на фиг. 10 D их только 2, но бывает и больше). Антеридии и у них образуют отроги, проникающие в оогонии, но переливания содержимого антеридия здесь не удалось наблюдать. Предполагают поэтому, что антеридии у сапролегний хотя и образуются, но более не функционируют. Это подкрепляется тем обстоятельством, что в некоторых случаях антеридиев и совсем не образуется, что не мешает яйцам в оогонии превращаться в нормальные ооспоры, которые происходят, значит, партеногенетически (партеноспоры). Итак, сапролегнии дают нам новый пример апогамии. Относительно существования О. у сумчатых грибов (Ascomycetes) мнения ботаников различны. Во всяком случае, где оно имеется, оно принадлежит к рассматриваемой категории гетерогамии. Женский орган, соответствующий оогонию, называется у них аскогоном (т. е. органом, производящим аскусы, сумки), а мужской, соответствующий антеридию пероноспоровых — поллинодием. Так как долго не могли заметить переливания содержимого из поллинодия в аскогон, а с другой стороны стали известны случаи образования сумок со спорами несомненно без всякого предварительного полового акта, то исследователи склонились считать все сумчатые грибы безвозвратно апогамными. Однако, недавно удалось Герперу (Harper) наблюдать, применяя метод окраски и микротомных разрезов, у сумчатого грибка Sphaerotheca слияние содержимого, а главное — слияние ядер мужского (поллинодия) с женским (аскогона). Оплодотворенный аскогон разрастается в толстую нить, состоящую из клеточек, из которых одна у Sphaerotheca скоро сильно вздувается и превращается в мешок (аскус) с 8 спорами. После этих исследований Герпера нужно принять, что у некоторых, по крайней мере, сумчатых грибов существует половой процесс. Гетерогамия этого же типа, но иногда в чрезвычайно усложненной форме существует у красных водорослей (см. Красные водоросли). Женский половой орган их — прокарп — состоит из двух частей: оплодотворяемого карпогона и передающей О. трихогины. Мужской орган — спермогоний (в нем образуются оплодотворяющие элементы) — мелкие шарообразные спермации. Содержимое спермаций, приносимых пассивно водой к трихогине, переливается через трихогину и сливается с яйцом, находящимся в карпогоне. В некоторых случаях констатировано и слияние мужского и женского ядер (Nemalion, Batrachospe r mum). Оплодотворенный карпогон образует сложным путем (см. Красные водоросли) карпоспоры.
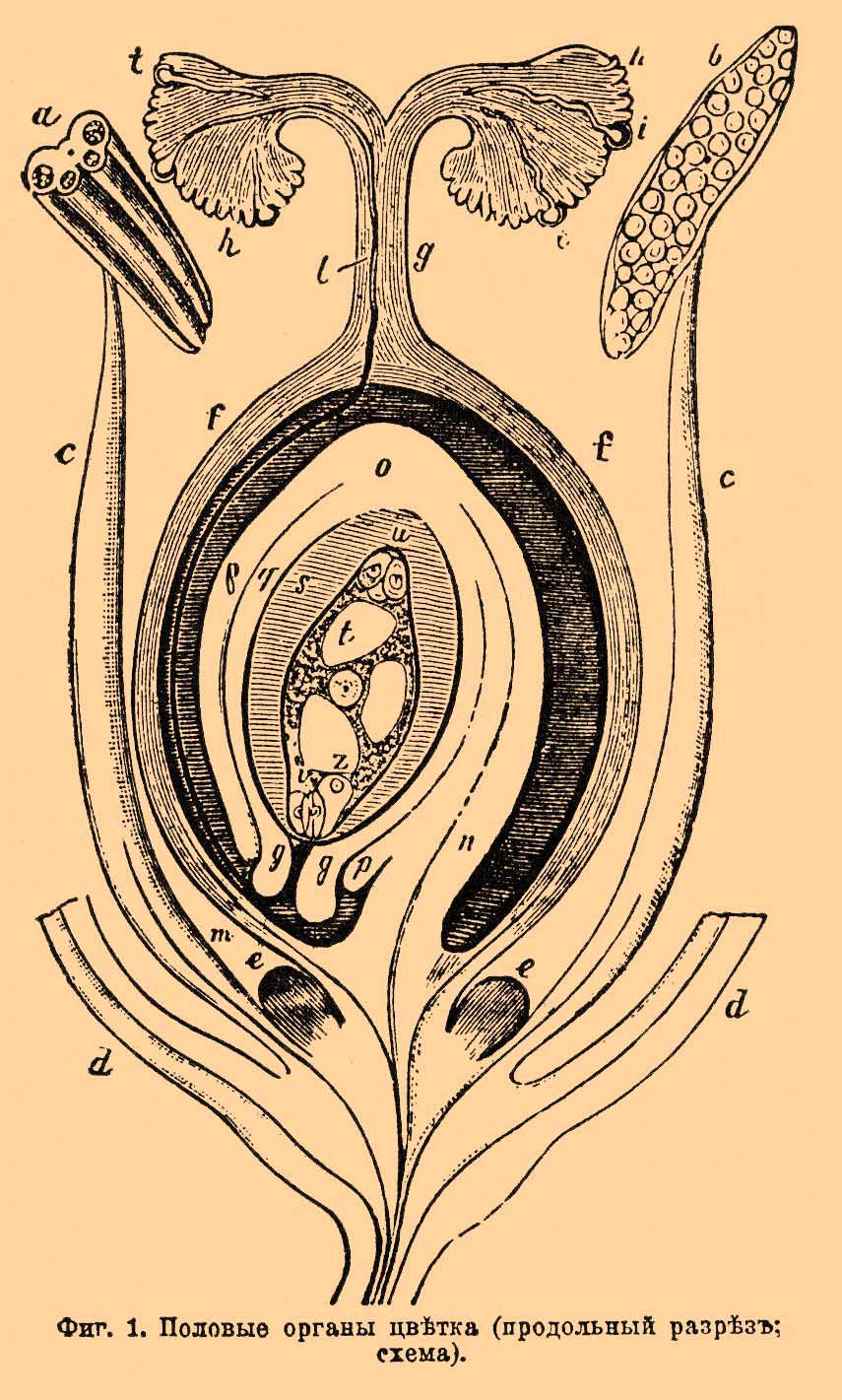
О. у высших (цветковых или семянных) растений происходит в форме гетерогамии. Мужские и женские половые клетки неспособны к активному передвижению, стало быть, апланогаметы; только очень недавно (1897) японские ученые Хиразе и Икено открыли существование сперматозоидов у двух цветковых растений — у саговой пальмы (Суcas revoluta) и у гинко (Ginkgo biloba); оба эти растения принадлежат к голосемянным (Gymnospermae), следовательно, к низшей группе среди цветковых растений. Познакомимся сначала с процессом О. у скрытосемянных (Angiospermae), к которым принадлежит огромное большинство цветковых растений. Мужские половые органы у них — тычинки, женские — пестики. Тычинка состоит из нити и прикрепленного к ее верхушке пыльника (см. схематическую фиг. 1, изображающую цветок [Познакомиться с устройством половых органов цветка всего лучше на крупных цветках, напр. лилии.] в разрезе; 2 тычинки, cc — их нити, а и b — пыльники; при а пыльник поперек перерезан, видны 4 гнезда, при b — вдоль, видны цветневые пылинки).
Фиг. 1. Половые органы цветка (продольный разрез; схема).
Внутри пыльника образуется цветень или иначе плодотворная пыльца; она состоит из мелких крупинок или пылинок (фиг. 1). Размеры и формы пылинок чрезвычайно разнообразны у различных растений (см. Цветень), для примера см. фиг. 2 и 3.
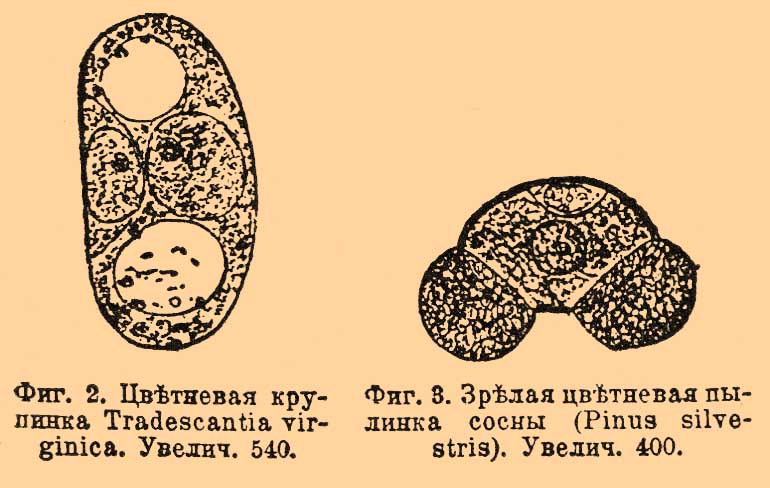
Фиг. 2. Цветневая крупинка Tradescantia virginica. Увелич. 540.
Фиг. 3. Зрелая цветневая пылинка сосны (Pinus silvestris). Увелич. 400.
Каждая пылинка одета двойной оболочкой — наружная, кутинизированная, называется экзиной, внутренняя, более нежная, состоящая преимущественно из пектиновых веществ — интиной. Поверхность пылинки часто покрыта шипиками, валиками или выпуклыми, нередко чрезвычайно красивыми узорами. Содержимое пылинки состоит из 2 голых клеточек очень неравной величины (см. фиг. 2). Маленькая клеточка называется антеридиальной, так как она соответствует антеридию высших споровых растений, а большая вегетативной — она соответствует всей остальной (вегетативной) части заростка у этих растений. Вся пылинка таким образом = микроспоре разноспоровых папоротников и плаунов (см. Водяные папоротники). Важно, что при О. играет роль только маленькая антеридиальная клеточка, вегетативная же прямого участия в этом процессе не принимает. Обращаясь снова к схематической фиг. 1 видим, что пестик состоит из 3 частей: завязи (f), столбика (g) и рыльца (h). Внутри завязи находится крупное яичко, называемое также семяпочкой, так как из нее происходит семя. Яичко сидит на ножке (n), посредством которой прикрепляется к нижней части (дну) завязи. Внутренняя часть яичка называется ядром (ss) — оно облечено 2 покровами (р и q; бывает и один покров). Место отхождения покровов от яичка получило наименование халазы (chalaza, о). Покровы не смыкаются вплотную, а оставляют небольшой узенький канал, ведущий вглубь яичка, это — семявход или микропиле (micropyle, т). Внутри яичка находится нежный зародышевый мешок (t), редко бывает несколько таких мешков. Заметим, что он соответствует макроспоре разноспоровых папоротников и плаунов. В зародышевом мешке посредине лежит клеточное ядро, а на концах по 3 голых (без оболочки) клеточки. Из них 3, лежащие ближе к семявходу, составляют так наз. лицевой аппарат, в состав которого входит яйцо (фиг. l, z; фиг. 4, о — при большем увеличении), которое отнюдь не нужно смешивать с вышеупомянутым яичком и 2 клеточки, остающиеся бесплодными (фиг. 1, v; фиг. 4, s) — их наз. синергидами или вспомогательными, хотя не известно с точностью, какую помощь оказывают они при О.

Фиг. 4. Funkia ovata. A — верхушка ядра яичка (nucellus, n) и зародышевого мешка с яйцевым аппаратом, до оплодотворения: o — яйцо, s — синергида. B — во время оплодотворения; t — пыльцевая трубочка. Увелич. 600.
Еще менее известно назначение 3 других клеточек, лежащих в противоположном конце мешка и называемых антиподами. Полагают, что все 6 клеток вместе представляют образование, соответствующее женскому заростку споровых растений, только в сильно редуцированном виде. Теперь обратимся к самому процессу О. Зрелая пыльца из пыльников переносится на рыльце пестика, у одних растений с помощью ветра, у других — воды, у третьих — насекомых (см. Опыление). Много пропадает при этом пыльцы, но некоторые пылинки достигают цели — происходит опыление. Попав на рыльце, пылинки прилипают к нему и под влиянием выделяемой рыльцем липкой жидкости — прорастают. При этом интина пылинки вытягивается в длинную пыльцевую или цветневую трубочку, нередко проходя через особые, заранее намеченные места в оболочке пылинки, — места, где экзина отсутствует или образует особые крышечки, приподнимаемые прорастающей трубочкой. Трубочка растет дальше и дальше, проходит сначала рыльце, потом столбик, спускаясь по особому канальцу столбика или пробираясь среди его центральной рыхлой паренхимы, раздвигая клетки, вступает затем в завязь и, наконец, достигает верхушки зародышевого мешка (фиг. 1, 1) . Обыкновенно прорастают несколько пылинок (фиг. 1, ii), но так как пыльцевые трубочки растут неодинаково быстро, то из них одна приходит первой к цели и производит О. Время, необходимое для прохождения трубки до яйца, иначе протекающее от опыления до О. — различно, иногда это всего несколько часов, иногда несколько дней, напр. у обыкновенного шафрана 3 дня, у орхидных 10 дней, а у сосны О. происходит лишь на другой год после опыления — пыльцевая трубочка у них замирает на зиму и продолжает расти следующей весной. Прорастание пыльцы может происходить не только на рыльце, но и в капле сахаристой жидкости или просто на ломтике арбуза и т. п. сладких плодов. Весьма вероятно (опыты японца Миуши), что пыльцевая трубочка стремится вглубь к зародышевому мешку под влиянием химического раздражения (хемотропизм пыльцевой трубочки), следовательно, под влиянием той же причины, которая заставляет живчиков у споровых растений стремиться внутрь архегония (ср. выше). Не всегда, однако, пыльцевая трубочка проникает к зародышевому мешку через естественный семявход. Исследования Трейба (Treub) показали, что у казуарин (см.) она подходит как раз с противоположного конца, именно через халазу. Трейб предложил даже отделить казуарин в особый класс — халазогамных растений, противопоставив их всем прочим скрытосемянным растениям — порогамным, у которых О. происходит через пору — семявход. Впоследствии исследования С. Г. Навашина показали, что халазогамия свойственна также некоторым самым обыкновенным у нас растениям — березе, ольхе, вязу и др. Тем или другим путем конец трубочки доходит до зародышевого мешка, оболочка мешка в этом месте растворяется и трубочка достигает, наконец, яйца. По мере роста трубочки передвигается и содержимое пылинки, так что самая пылинка и более старые части трубочки постепенно опоражниваются. Во время передвижения антеридиальная клеточка пылинки делится на 2 голых генеративных клеточки. На фиг. 5 А изображен конец пыльцевой трубочки с 2 генеративными клеточками (gz, gz,) впереди их ядро вегетативной клетки (ok), которая собственно и вытянулась в трубочку, увлекая с собой антеридиальную клетку.

Фиг. 5. Процесс оплодотворения у цветкового скрытосемянного растения (несколько схематично). Увелич. около 500.
Ядро это разрушается и никакого участия в О. не принимает. Оплодотворяет одна из генеративных клеточек: она выскользает из трубочки в яйцо и сливается с ним. Та же фиг. 5, B, C, D представляет главные моменты О. В — проникновение в яйцо генеративной клеточки, ядру клеточки (sk) предшествуют 2 центрозомы (с), у ядра яйца (ek) также пара центрозом (с), ch — зачатки хроматофоров. С — начало слияния ядра генеративной клеточки (мужского) с ядром яйца (женским). D — продукт слияния, оплодотворенное яйцо, мужское и женское ядра слились в зачатковое ядро (kk). Протоплазма генеративной клеточки сливается также с протоплазмой яйца, равным образом сливаются (по Гиньяру) и их центрозомы и дают пару центрозом зачаткового ядра (cc). По другим показаниям, центрозомы зачаткового ядра ведут свое происхождение от центрозом яйца или, гораздо реже, от центрозом генеративной клетки. Что касается до хроматофоров, то их поставляет для будущего растения одно яйцо, без участия мужской клетки. Оплодотворенное яйцо окружается тонкой оболочкой, потом делится и превращается в зародыш. Что касается до синергид, то они скоро разрушаются и исчезают; по-видимому, содержимое их идет на питание оплодотворенного яйца. Особый интерес представляет судьба хроматина клеточных ядер при процессе О., прослеженная всего лучше у лилии. Обыкновенные ядра вегетативных клеток у этого растения дают при кариокинетическом делении 24 хроматиновых сегмента (хромосомы), которые, расщепляясь вдоль пополам, в том же числе 24-х переходят в дочерние ядра. Когда же образуются половые клетки, происходит сокращение — редукция хромосом, так что в клеточных ядрах яйца и генеративной клеточки оказывается вдвое меньше хромосом, т. е. по 12 в каждом. Благодаря этому обстоятельству при слиянии мужского ядра с женским получается зачатковое ядро с нормальным количеством хромосом, из коих 12 отцовского и 12 материнского происхождения. — Перейдем теперь к голосемянным. Они отличаются некоторыми существенными чертами в устройстве половых органов и прежде всего тем, что у них нет завязи. Яичко у них состоит из ядра (nucellus) — фиг. 6, nc, окруженного одним покровом (i); семявход очень широк, так что цветневые пылинки (р) попадают прямо на ядро яичка и потому цветневые трубочки (t) у голосемянных гораздо короче, нежели у скрытосемянных, что не мешает им у сосны, как упомянуто выше, очень долго пробираться до яйца.
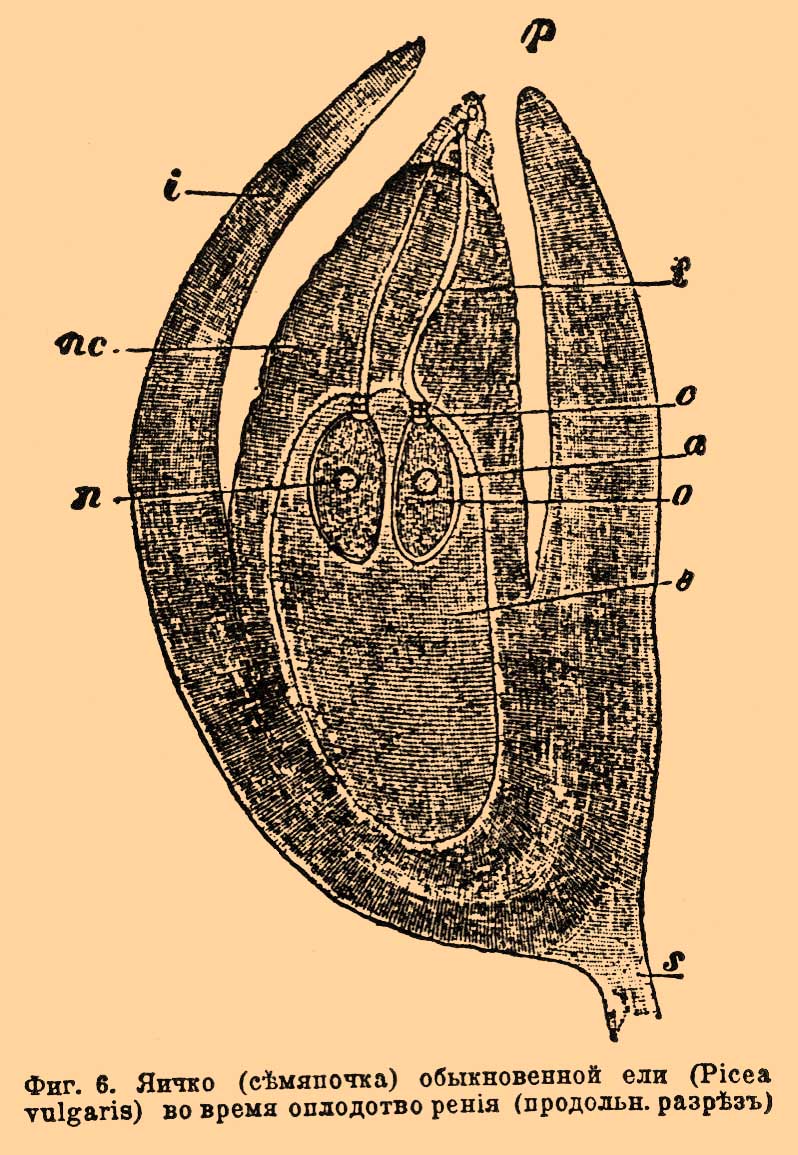
Фиг. 6. Яичко (семяпочка) обыкновенной ели (Picea vulgaris) во время оплодотворения (продольн. разрез).
В ядре находится зародышевый мешок, как и у скрытосемянных, но устройство его иное и он весь заполняется еще до О. паренхиматическими клетками. Некоторые из этих клеток, расположенные на верхушке мешка, превращаются в так назыв. корпускулы или архегонии — органы совершенно соответствующие по строению и по назначению архегониям высших споровых растений; их бывает от 2 до 15 (см. фиг. 6, где их представлено 2) [Весь комплекс клеток, заполняющий зародышевый мешок, соответствует женскому заростку, а весь мешок целиком, как и у скрытосемянных — макроспоре; микроспоре соответствует и здесь цветневая пылинка]. В основной вздутой части архегонии лежит яйцо (о). Содержимое цветневой пылинки дифференцируется у голосемянных также на 2 неравные клеточки, но они не голые, так как между ними находится перегородка. Фиг. 3 изображает пылинку сосны; форма ее весьма характерна — пылинка имеет по бокам два пузыря, наполненных воздухом — они произошли через выпячивание экзины; пузыри эти облегчают перенесение пылинки ветром. Скоро внутри пылинки оказывается не две, а несколько маленьких клеточек, покрытых оболочками, из них одна становится антеридиальной. При прорастании пылинки антеридиальная клеточка перемещается в конец трубочки и распадается на 2 генеративных клеточки. О. заключается, как и у скрытосемянных, в слиянии генеративной клеточки с яйцом. Фиг. 7 А, B, С представляет постепенные стадии процесса О. у обыкновенной ели: клеточное ядро генеративной клеточки (sn) выскальзывает из конца пыльцевой трубочки (p), подходит к ядру яйца (on) и сливается с ним.
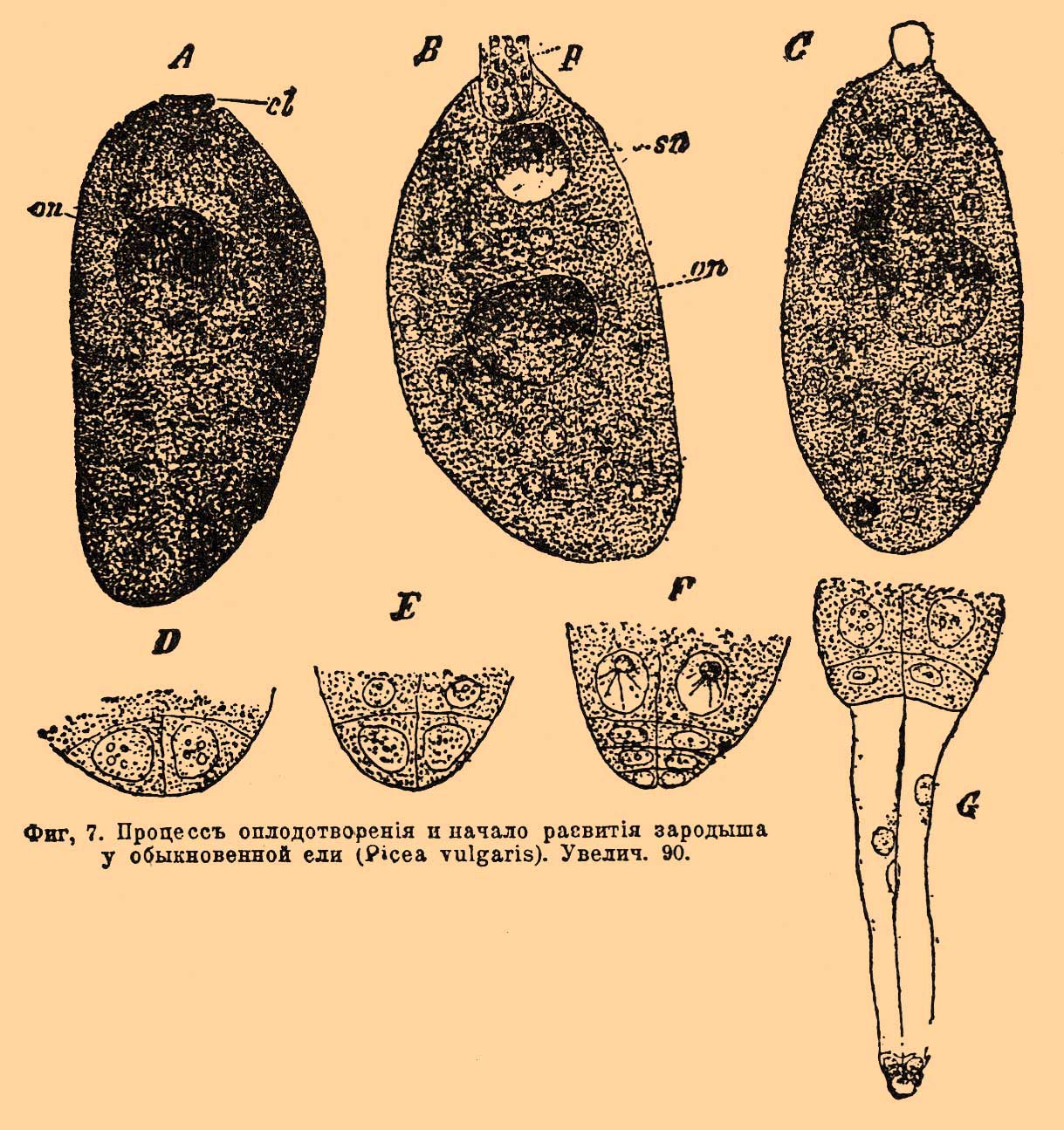
Фиг. 7. Процесс оплодотворения и начало развития зародыша у обыкновенной ели (Picea vulgaris). Увелич. 90.
Произошедшее через слияние зачатковое ядро спускается на дно яйца (вместе с тем и архегониия) и здесь повторным делением дает 4 ядра; скоро мы находим в этом месте 4 клеточки, расположенные в одной плоскости (фиг. 7, D). Клеточки делятся поперечными перегородками, так что получается несколько четырехклетных этажей (E, F). Затем, один из этажей вытягивается в 4 длинные нити, унося на вершине вглубь заростка клетки, служащие для развития зародыша (G). Таким образом, начинают формироваться 4 зародыша, но доразвивается обыкновенно только один. Равным образом, хотя было бы оплодотворенно и несколько архегониев, в зрелом семени находится один только зародыш, как и у скрытосемянных. Зародыш этот лежит среди остатков заростка или, как его называют здесь — эндосперма (белка). Так как описанный способ передачи мужского элемента О. через трубочку (пыльцевую) характерен для всех цветковых растений и отличает их от всех высших споровых, то Энглер предложил называть эти растения сифоногамными (Embryophyta siphonogama), противопоставляя их производящим сперматозоиды высшим споровым — зоидиогамным растениям (Zoidiogamae). Недавно, однако, Хиразе и Икено открыли существование сперматозоидов и у цветковых растений — у Cycas и Ginkgo, принадлежащих к голосемянным. В конце пыльцевой трубочки у них генеративные клеточки превращаются в 2 овальных сперматозоида, снабженных многими ресничками. Сперматозоиды плавают довольно быстро в жидкости, окружающей архегонии, добираются до яйца и оплодотворяют его. Причина образования сперматозоидов кроется, по мнению Икено, в том, что пыльцевая трубочка у этих растений очень коротка и не доходит до архегония, так что без помощи подвижных мужских гамет, т. е. сперматозоидов, О. у них было бы невозможно. Во всяком случае, существуют растения одновременно сифоногамные и зоидиогамные, являющиеся в высшей степени интересной переходной формой по своему О. между высшими споровыми и цветковыми растениями. Специальная литература об О. у растений чрезвычайно обширна. Подробности и литературные цитаты можно найти в следующих сочинениях: проф. И. П. Бородин, "Процесс оплодотворения в растительном царстве" (2-е изд., 1896); F. Pax, "Allgemeine Morphologie der Pflanzen" (1890); van-Tieghem, "Trait é de botanique", том I , 1891; E. Strasburger, "Das botanische Practicum" (3 изд., 18 97); G. Klebs, "Die Bedingungen der Fortpflanzung bei einigen Algen und Pilzen" (1896); S. Hirase, "Untersuchungen ü ber das Verhalten des Pollens von Ginkgo biloba" ("Botanisches Centralblatt", 1897, № 2—3); prof. I. Ikeno, "Vorla üfige Mittheilung über die Spermatozoiden bei Cycas revoluta" (там же, № 1).
Г. Надсон.